

Specific antibodies to soluble alpha-synuclein conformations in intravenous immunoglobulin preparations
- PMID: 20646004
- PMCID: PMC2962971
- DOI: 10.1111/j.1365-2249.2010.04214.x
Specific antibodies to soluble alpha-synuclein conformations in intravenous immunoglobulin preparations
Abstract
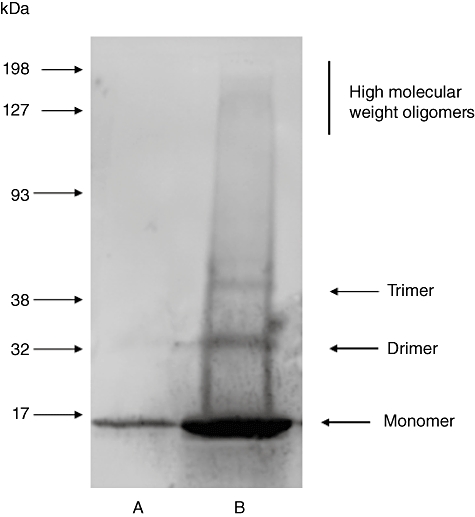
Alpha-synuclein is the major protein in Lewy bodies, the hallmark pathological finding in Parkinson's disease (PD) and dementia with Lewy bodies (DLB). Although normally intracellular, it also can be secreted, so extracellular alpha-synuclein may contribute to neuronal injury. Serum antibodies to alpha-synuclein could exert protective effects by increasing alpha-synuclein's movement out of the brain and, if they cross the blood-brain barrier, by inhibiting its neurotoxic effects. The objective of this study was to measure antibody concentrations to alpha-synuclein monomer and soluble oligomers in three intravenous immunoglobulin (IVIG) preparations, Gamunex (Talecris Biotherapeutics), Gammagard (Baxter Healthcare) and Flebogamma (Grifols Biologicals). Antibodies were measured in native IVIG preparations and after antibody-antigen complex dissociation. IVIG's non-specific binding was subtracted from its total binding to alpha-synuclein to calculate specific anti-alpha-synuclein antibody concentrations. Specific antibodies to alpha-synuclein monomer and/or soluble oligomers were detected in all IVIG products. In native IVIG preparations, the highest anti-monomer concentrations were in Gammagard and the highest anti-oligomer concentrations were in Gamunex; the extent to which lot-to-lot variation may have contributed to these differences was not determined. Antibody-antigen complex dissociation had variable effects on these antibody levels. The IVIG preparations did not inhibit alpha-synuclein oligomer formation, although they changed the distribution and intensity of some oligomer bands on Western blots. The presence of antibodies to soluble alpha-synuclein conformations in IVIG preparations suggests that their effects should be studied in animal models of synucleinopathies, as a first step to determine their feasibility as a possible treatment for PD and other synucleinopathies.
© 2010 British Society for Immunology.
Figures





Similar articles
-
Antibody concentrations to Abeta1-42 monomer and soluble oligomers in untreated and antibody-antigen-dissociated intravenous immunoglobulin preparations.Int Immunopharmacol. 2010 Jan;10(1):115-9. doi: 10.1016/j.intimp.2009.10.005. Epub 2009 Oct 17. Int Immunopharmacol. 2010. PMID: 19840873
-
Effects of intravenous immunoglobulin on alpha synuclein aggregation and neurotoxicity.Int Immunopharmacol. 2012 Dec;14(4):550-7. doi: 10.1016/j.intimp.2012.09.007. Epub 2012 Sep 29. Int Immunopharmacol. 2012. PMID: 23026444
-
Intravenous immunoglobulin products contain specific antibodies to recombinant human tau protein.Int Immunopharmacol. 2013 Aug;16(4):424-8. doi: 10.1016/j.intimp.2013.04.034. Epub 2013 May 10. Int Immunopharmacol. 2013. PMID: 23669333
-
Antibodies against alpha-synuclein: tools and therapies.J Neurochem. 2019 Sep;150(5):612-625. doi: 10.1111/jnc.14713. Epub 2019 Jun 25. J Neurochem. 2019. PMID: 31055836 Review.
-
The therapeutical potential of alpha-synuclein antiaggregatory agents for dementia with Lewy bodies.Curr Med Chem. 2008;15(26):2748-59. doi: 10.2174/092986708786242868. Curr Med Chem. 2008. PMID: 18991634 Review.
Cited by
-
Intravenous immunoglobulin and Alzheimer's disease: what now?J Neuroinflammation. 2013 Jun 5;10:70. doi: 10.1186/1742-2094-10-70. J Neuroinflammation. 2013. PMID: 23735288 Free PMC article. Review.
-
α-Synuclein and anti-α-synuclein antibodies in Parkinson's disease, atypical Parkinson syndromes, REM sleep behavior disorder, and healthy controls.PLoS One. 2012;7(12):e52285. doi: 10.1371/journal.pone.0052285. Epub 2012 Dec 17. PLoS One. 2012. PMID: 23284971 Free PMC article.
-
Alzheimers disease: review of emerging treatment role for intravenous immunoglobulins.J Cent Nerv Syst Dis. 2011 May 8;3:67-73. doi: 10.4137/JCNSD.S5018. Print 2011. J Cent Nerv Syst Dis. 2011. PMID: 23861639 Free PMC article.
-
Human serum-derived α-synuclein auto-antibodies mediate NMDA receptor-dependent degeneration of CNS neurons.J Neuroinflammation. 2024 Feb 28;21(1):62. doi: 10.1186/s12974-024-03050-6. J Neuroinflammation. 2024. PMID: 38419079 Free PMC article.
-
Novel therapeutic approaches in multiple system atrophy.Clin Auton Res. 2015 Feb;25(1):37-45. doi: 10.1007/s10286-014-0249-7. Epub 2014 Jun 14. Clin Auton Res. 2015. PMID: 24928797 Free PMC article. Review.
References
-
- Giasson BI, Murray IV, Trojanowski JQ, Lee VM. A hydrophobic stretch of 12 amino acid residues in the middle of alpha-synuclein is essential for filament assembly. J Biol Chem. 2001;276:2380–6. - PubMed
-
- Uversky VN, Lee HJ, Li J, Fink AL, Lee SJ. Stabilization of partially folded conformation during alpha-synuclein oligomerization in both purified and cytosolic preparations. J Biol Chem. 2001;276:43495–8. - PubMed
-
- Miake H, Mizusawa H, Iwatsubo T, Hasegawa M. Biochemical characterization of the core structure of alpha-synuclein filaments. J Biol Chem. 2002;277:19213–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
