

Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis
- PMID: 20647342
- PMCID: PMC2929093
- DOI: 10.1105/tpc.110.076588
Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that requires EIN2 in Arabidopsis
Abstract
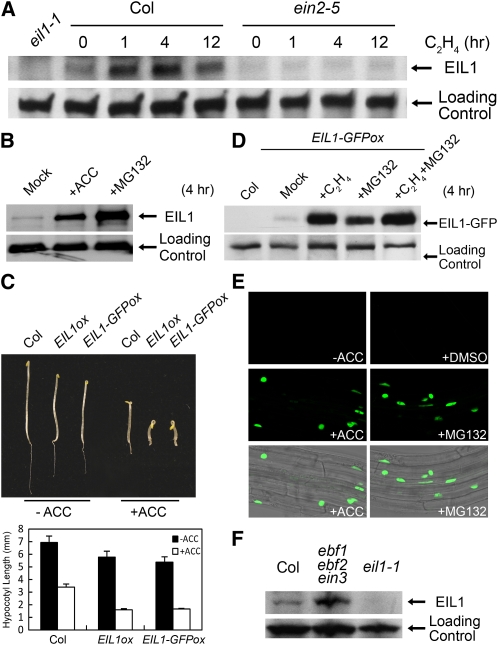
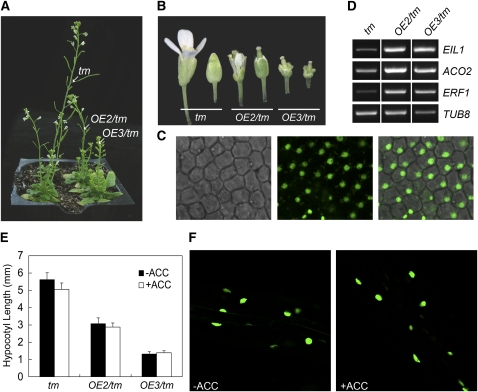
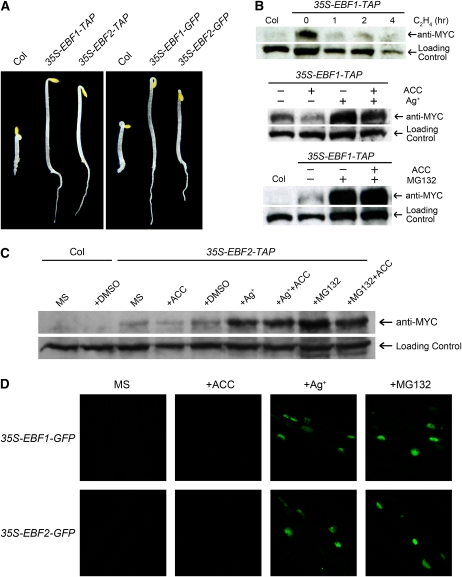
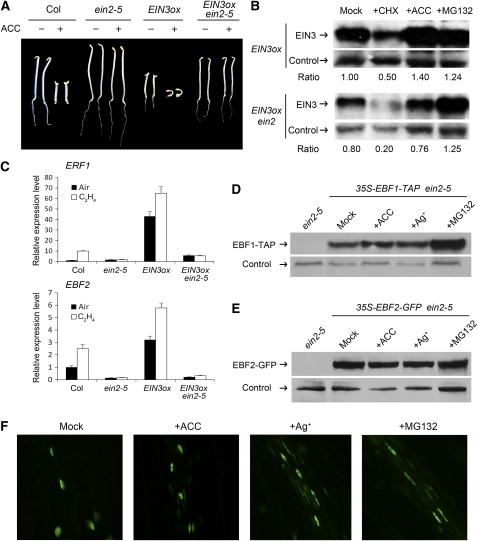
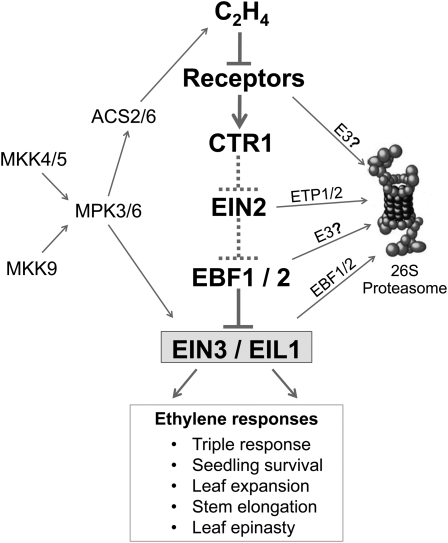
Plant responses to ethylene are mediated by regulation of EBF1/2-dependent degradation of the ETHYLENE INSENSITIVE3 (EIN3) transcription factor. Here, we report that the level of EIL1 protein is upregulated by ethylene through an EBF1/2-dependent pathway. Genetic analysis revealed that EIL1 and EIN3 cooperatively but differentially regulate a wide array of ethylene responses, with EIL1 mainly inhibiting leaf expansion and stem elongation in adult plants and EIN3 largely regulating a multitude of ethylene responses in seedlings. When EBF1 and EBF2 are disrupted, EIL1 and EIN3 constitutively accumulate in the nucleus and remain unresponsive to exogenous ethylene application. Further study revealed that the levels of EBF1 and EBF2 proteins are downregulated by ethylene and upregulated by silver ion and MG132, suggesting that ethylene stabilizes EIN3/EIL1 by promoting EBF1 and EBF2 proteasomal degradation. Also, we found that EIN2 is indispensable for mediating ethylene-induced EIN3/EIL1 accumulation and EBF1/2 degradation, whereas MKK9 is not required for ethylene signal transduction, contrary to a previous report. Together, our studies demonstrate that ethylene similarly regulates EIN3 and EIL1, the two master transcription factors coordinating myriad ethylene responses, and clarify that EIN2 but not MKK9 is required for ethylene-induced EIN3/EIL1 stabilization. Our results also reveal that EBF1 and EBF2 act as essential ethylene signal transducers that by themselves are subject to proteasomal degradation.
Figures





References
-
- Abeles F.B., Morgan P.W., Saltveit J.M.E. (1992). Ethylene in Plant Biology. 2nd ed (San Diego, CA: Academic Press; ).
-
- Alonso J.M., Hirayama T., Roman G., Nourizadeh S., Ecker J.R. (1999). EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
-
- Alonso J.M., et al. (2003b). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Bechtold N., Pelletier G. (1998). In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 82: 259–266 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
