

Repair of mitomycin C mono- and interstrand cross-linked DNA adducts by UvrABC: a new model
- PMID: 20647419
- PMCID: PMC2978355
- DOI: 10.1093/nar/gkq576
Repair of mitomycin C mono- and interstrand cross-linked DNA adducts by UvrABC: a new model
Abstract
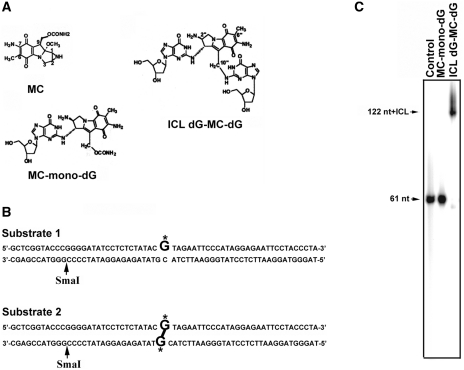
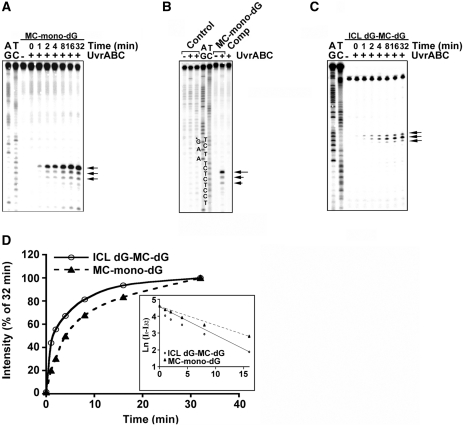
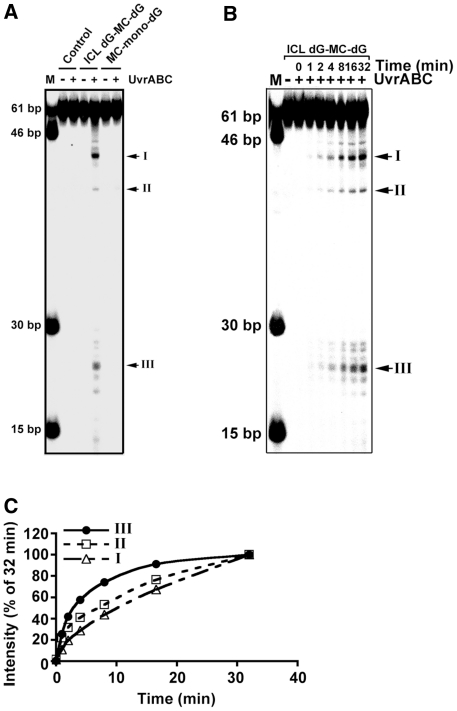
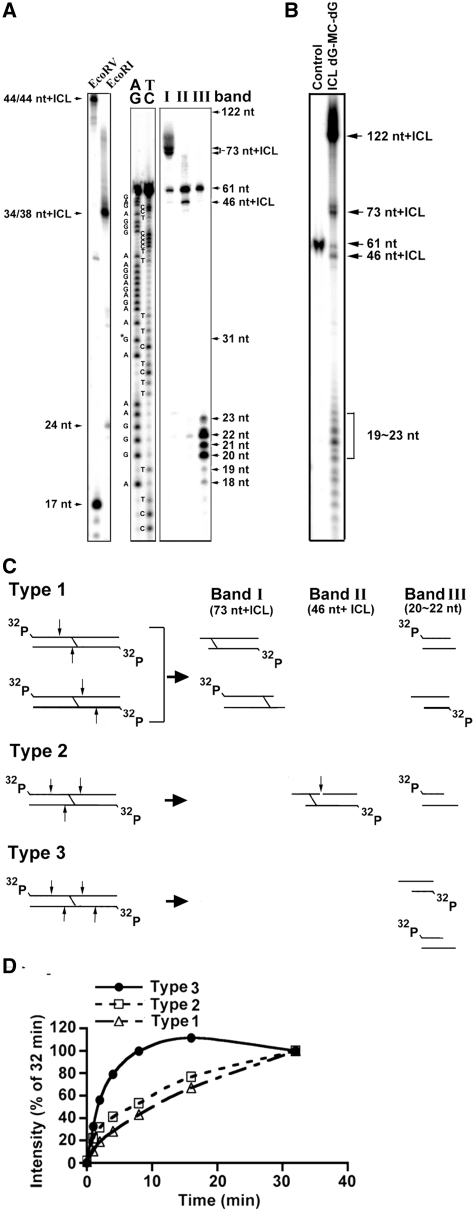
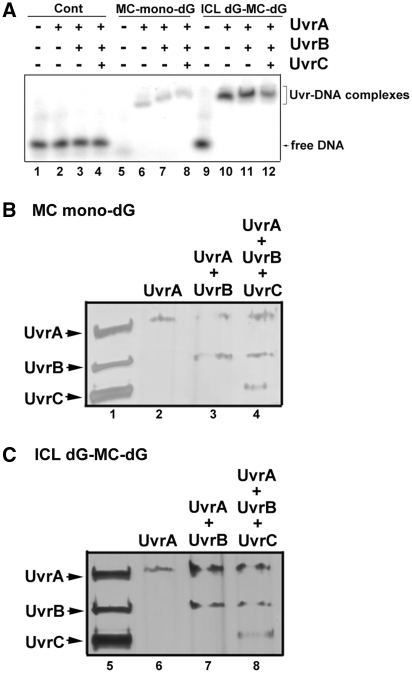
Mitomycin C induces both MC-mono-dG and cross-linked dG-adducts in vivo. Interstrand cross-linked (ICL) dG-MC-dG-DNA adducts can prevent strand separation. In Escherichia coli cells, UvrABC repairs ICL lesions that cause DNA bending. The mechanisms and consequences of NER of ICL dG-MC-dG lesions that do not induce DNA bending remain unclear. Using DNA fragments containing a MC-mono-dG or an ICL dG-MC-dG adduct, we found (i) UvrABC incises only at the strand containing MC-mono-dG adducts; (ii) UvrABC makes three types of incisions on an ICL dG-MC-dG adduct: type 1, a single 5' incision on 1 strand and a 3' incision on the other; type 2, dual incisions on 1 strand and a single incision on the other; and type 3, dual incisions on both strands; and (iii) the cutting kinetics of type 3 is significantly faster than type 1 and type 2, and all of 3 types of cutting result in producing DSB. We found that UvrA, UvrA+UvrB and UvrA+UvrB+UvrC bind to MC-modified DNA specifically, and we did not detect any UvrB- and UvrB+UvrC-DNA complexes. Our findings challenge the current UvrABC incision model. We propose that DSBs resulted from NER of ICL dG-MC-dG adducts contribute to MC antitumor activity and mutations.
Figures
References
-
- Kusenda Z, Kerger J, Awada A, Geurs F, Van Vreckem A, Habboubi N, Piccart MJ. Mitomycin C and vinblastine in combination with amifostine in metastatic breast cancer. A feasibility study of the EORTC–Investigational Drug Branch for Breast Cancer (IDBBC) Support Care Cancer. 1997;5:414–416. - PubMed
-
- Lenaz L. Mitomycin C in advanced breast cancer. Cancer Treat. Rev. 1985;12:235–249. - PubMed
-
- Bolenz C, Cao Y, Arancibia MF, Trojan L, Alken P, Michel MS. Intravesical mitomycin C for superficial transitional cell carcinoma. Expert Rev. Anticancer Ther. 2006;6:1273–1282. - PubMed
-
- Snegovoy A, Manzuk L, Artamonova E. Mitomycin plus capecitabine in metastatic breast cancer. 2008 Breast Cancer Symposium. 2008 General Poster Session E, Abstract No: 264.
-
- Bizanek R, McGuinness BF, Nakanishi K, Tomasz M. Isolation and structure of an intrastrand cross-link adduct of mitomycin C and DNA. Biochemistry. 1992;31:3084–3091. - PubMed
