

Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina
- PMID: 20648062
- PMCID: PMC11346175
- DOI: 10.1038/nrn2880
Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina
Abstract
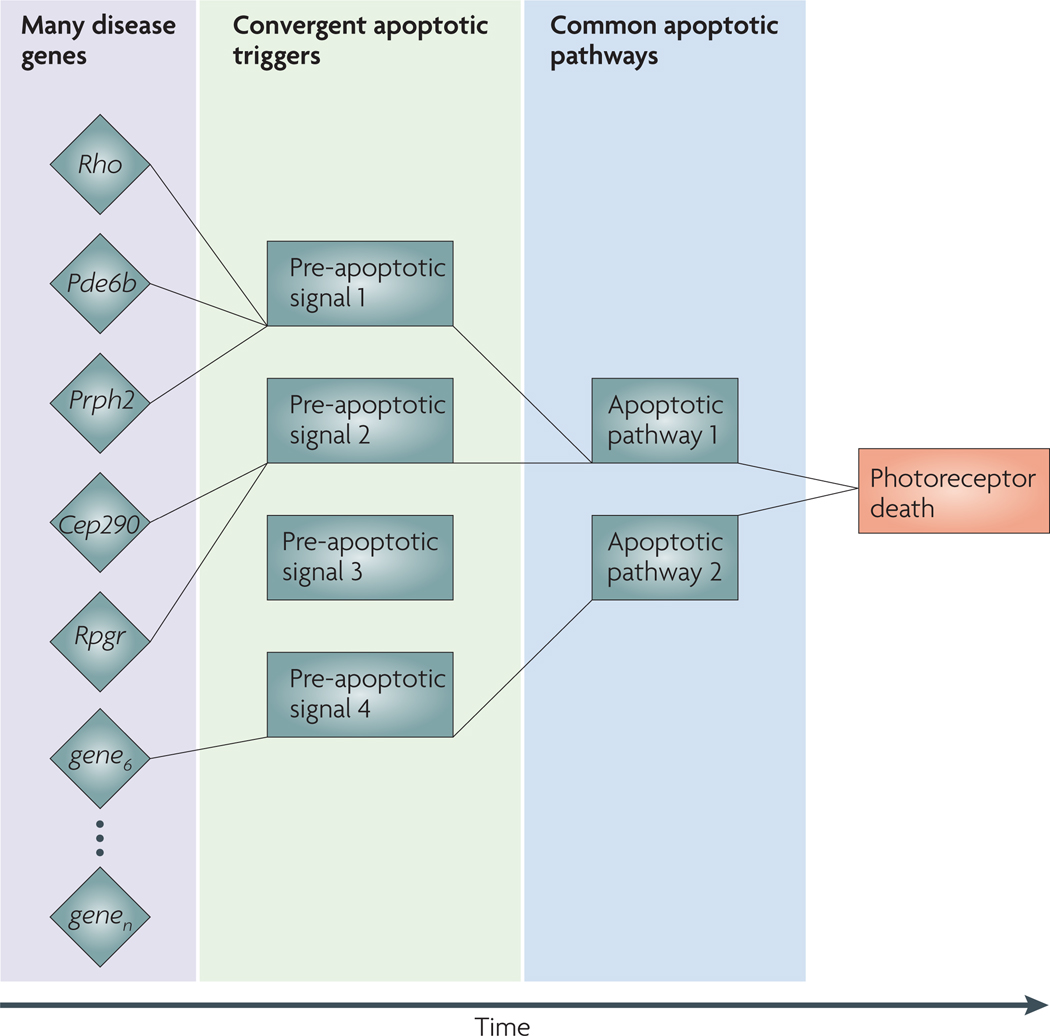
In the developing vertebrate retina, diverse neuronal subtypes originate from multipotent progenitors in a conserved order and are integrated into an intricate laminated architecture. Recent progress in mammalian photoreceptor development has identified a complex relationship between six key transcription-regulatory factors (RORbeta, OTX2, NRL, CRX, NR2E3 and TRbeta2) that determine rod versus M cone or S cone cell fate. We propose a step-wise 'transcriptional dominance' model of photoreceptor cell fate determination, with the S cone representing the default state of a generic photoreceptor precursor. Elucidation of gene-regulatory networks that dictate photoreceptor genesis and homeostasis will have wider implications for understanding the development of nervous system function and for the treatment of neurodegenerative diseases.
Conflict of interest statement
Competing interests statement
The authors declare no competing financial interests.
Figures
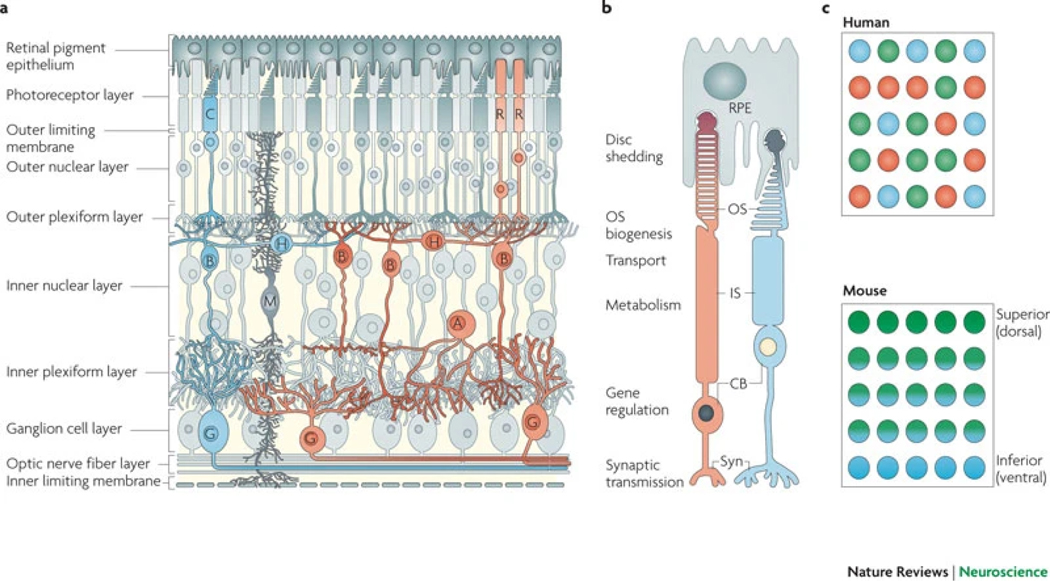
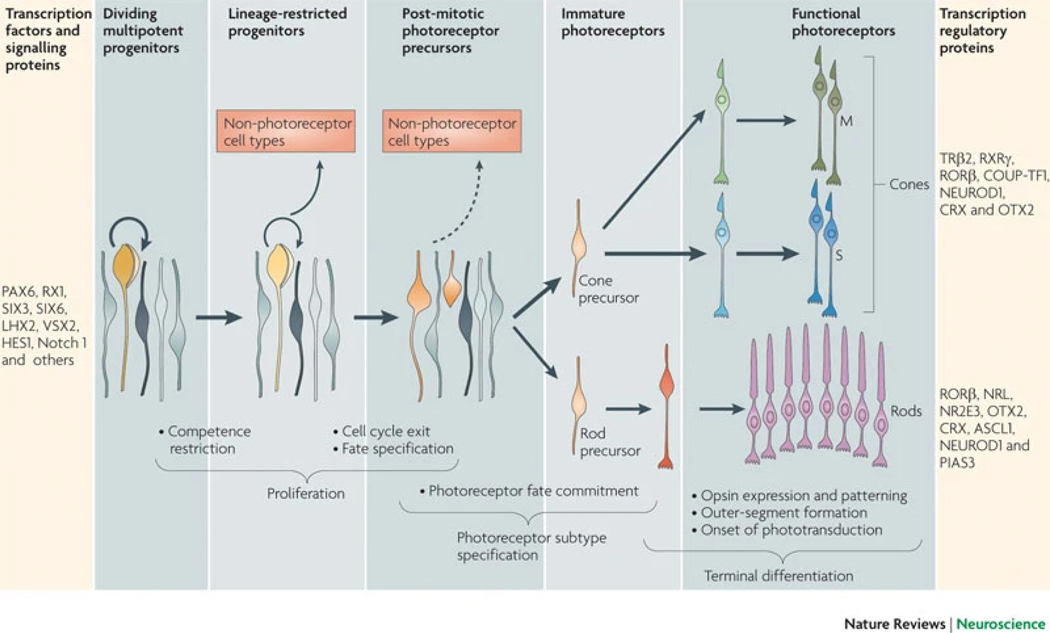
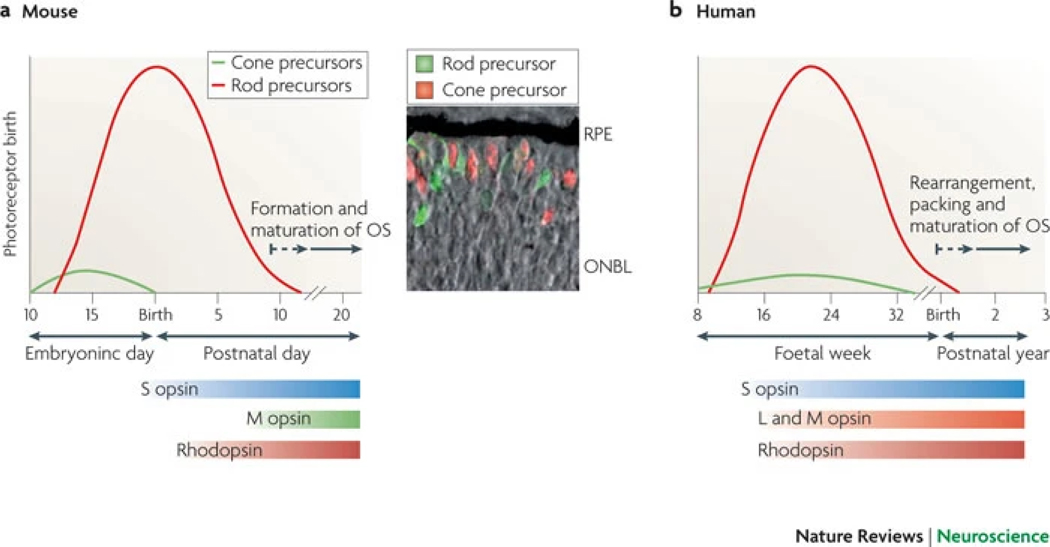

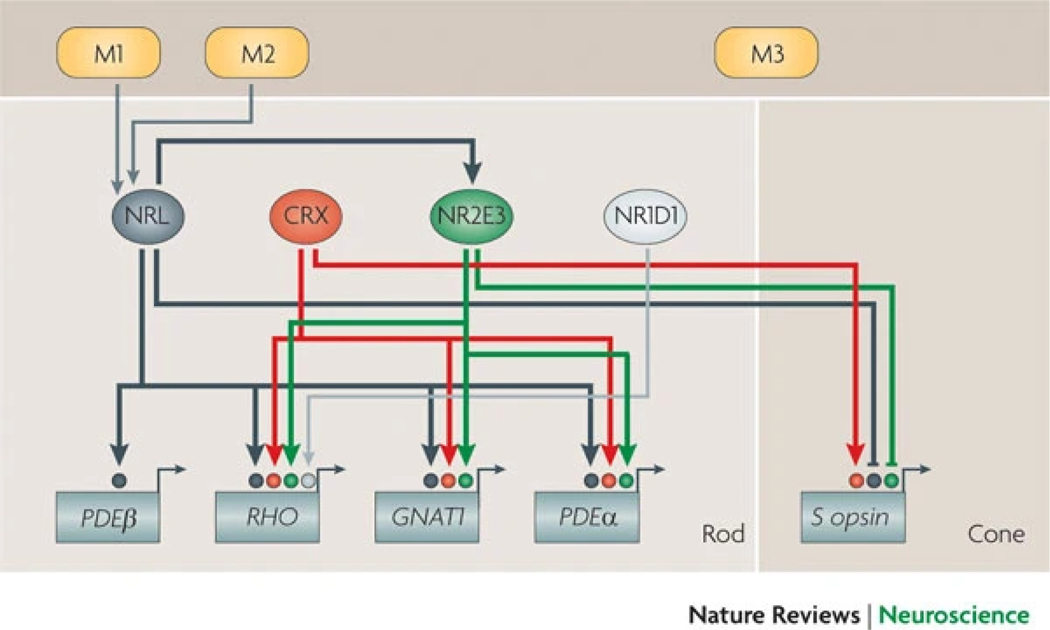
References
-
- Rodieck RW The First Steps in Seeing (Sinauer Associates Publishers, Sunderland, Massachusetts,1998).
-
- Dowling JE The Retina: An Approachable Part of the Brain. (Belknap Press, Harvard Univ. Press, 1987).
-
- Masland RH The fundamental plan of the retina. Nature Neurosci. 4, 877–886 (2001). - PubMed
-
- Wassle H. Parallel processing in the mammalian retina. Nature Rev. Neurosci. 5, 747–757 (2004). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
