

Bioenergetics and cell death
- PMID: 20649539
- PMCID: PMC3079367
- DOI: 10.1111/j.1749-6632.2010.05633.x
Bioenergetics and cell death
Abstract
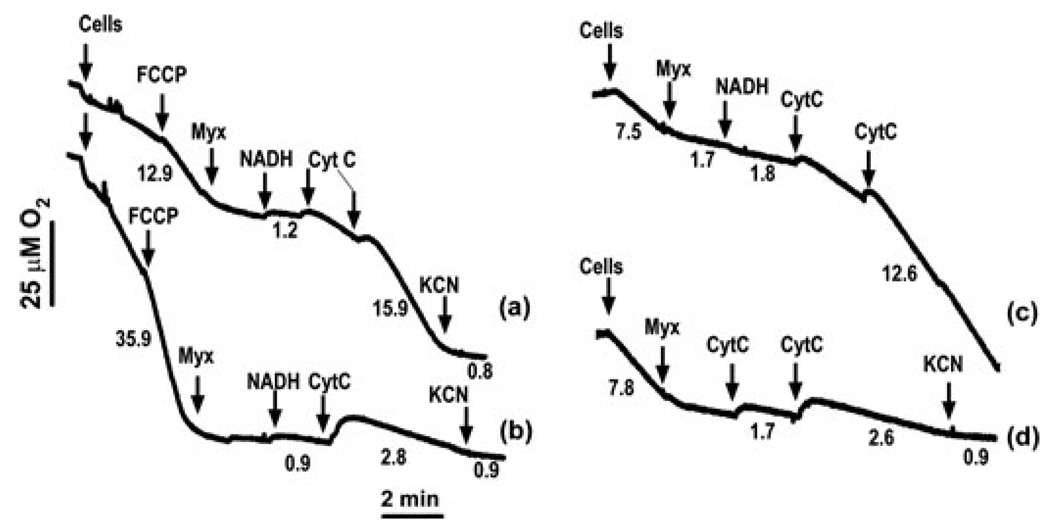
Mitochondrial bioenergetic function is a key to cell life and death. Cells need energy not only to support their vital functions but also to die gracefully. Execution of an apoptotic program includes energy-dependent steps, including kinase signaling, formation of the apoptosome, and effector caspase activation. Under conditions of bioenergetic collapse, cells are diverted toward necrotic demise. Mitochondrial outer membrane permeabilization (MOMP) is a decisive event in the execution of apoptosis. It is also causally linked to a decline in bioenergetic function via different mechanisms, not merely due to cytochrome c dispersion. MOMP-induced bioenergetic deficiency is usually irreversible and commits cells to die, even when caspases are inactive. Here, we discuss the mechanisms by which MOMP impacts bioenergetics in different cell death paradigms.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ricci JE, et al. Disruption of mitochondrial function during apoptosis is mediated by caspase cleavage of the p75 subunit of complex I of the electron transport chain. Cell. 2004;117:773–786. - PubMed
-
- Ankarcrona M, et al. Glutamate-induced neuronal death: a succession of necrosis or apoptosis depending on mitochondrial function. Neuron. 1995;15:961–973. - PubMed
-
- Nicotera P, Melino G. Regulation of the apoptosis-necrosis switch. Oncogene. 2004;23:2757–2765. - PubMed
-
- Skulachev VP. Bioenergetic aspects of apoptosis, necrosis and mitoptosis. Apoptosis. 2006;11:473–485. - PubMed
-
- Colell A, et al. GAPDH and autophagy preserve survival after apoptotic cytochrome c release in the absence of caspase activation. Cell. 2007;129:983–997. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
