

Resveratrol inhibits renal fibrosis in the obstructed kidney: potential role in deacetylation of Smad3
- PMID: 20651248
- PMCID: PMC2928940
- DOI: 10.2353/ajpath.2010.090923
Resveratrol inhibits renal fibrosis in the obstructed kidney: potential role in deacetylation of Smad3
Abstract
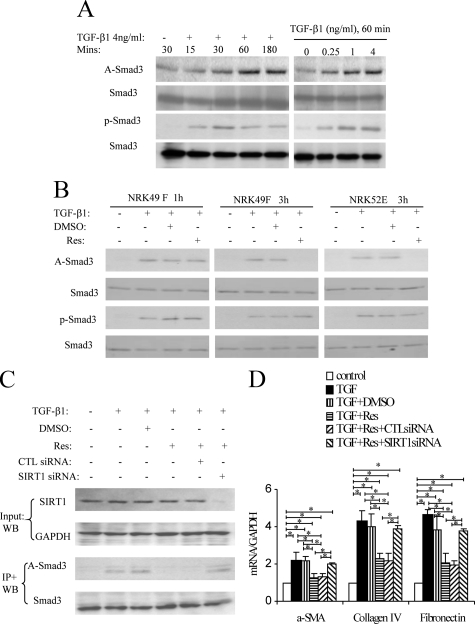
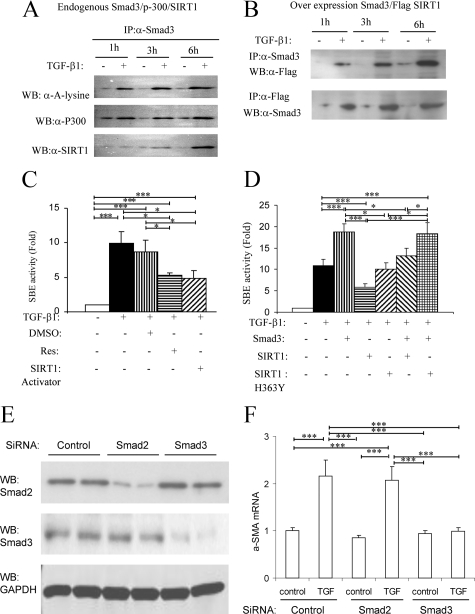
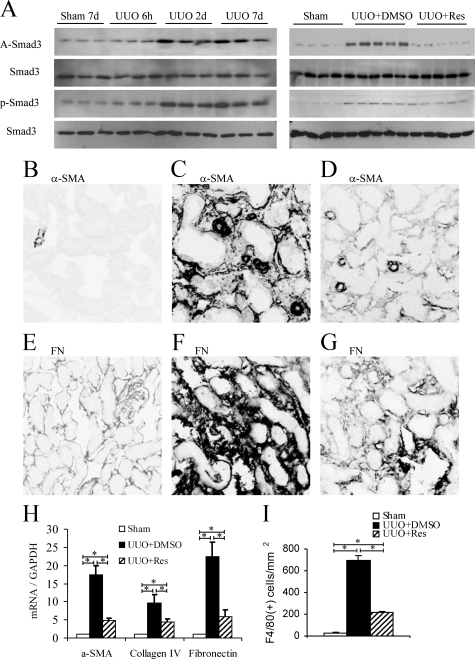
Transforming growth factor-beta1 (TGF-beta1) promotes tissue fibrosis through the Smad3 signaling pathway. While phosphorylation is known to regulate Smad3 function, recent in vitro studies have suggested that acetylation may also regulate Smad3 function. This study investigated Smad3 acetylation in renal fibrosis. TGF-beta1 stimulation of renal fibroblasts and tubular epithelial cells induced Smad3 acetylation and phosphorylation. Resveratrol, an activator of the Nicotinamide adenine dinucleotide (NAD) dependent protein deacetylase SIRT1, reversed acetylation but not phosphorylation of Smad3 and inhibited TGF-beta1-induced up-regulation of collagen IV and fibronectin mRNA levels. Knockdown of SIRT1 expression abolished the inhibitory effect of resveratrol, and co-immunoprecipitation studies provide direct evidence of an interaction between acetylated Smad3 and SIRT1. The role of Smad3 acetylation in renal fibrosis was then examined in the unilateral ureteric obstruction (UUO) model. Immunoprecipitation studies showed acetylation and phosphorylation of Smad3 by day 2 UUO, which was sustained to day 7 in association with development of interstitial fibrosis. Resveratrol inhibited acetylation but not phosphorylation of Smad3 at day 2 UUO, and resveratrol treatment inhibited interstitial fibrosis at day 7 UUO. In conclusion, these studies support a pathological role for Smad3 acetylation in renal fibrosis and suggest that deacetylation of Smad3 may be a novel therapeutic target for fibrotic disease.
Figures
References
-
- Wang W, Koka V, Lan HY. Transforming growth factor-beta and Smad signalling in kidney diseases. Nephrology. 2005;10:48–56. - PubMed
-
- Fujimoto M, Maezawa Y, Yokote K, Joh K, Kobayashi K, Kawamura H, Nishimura M, Roberts AB, Saito Y, Mori S. Mice lacking Smad3 are protected against streptozotocin-induced diabetic glomerulopathy. Biochem Biophys Res Commun. 2003;305:1002–1007. - PubMed
-
- Li JH, Huang XR, Zhu HJ, Oldfield M, Cooper M, Truong LD, Johnson RJ, Lan HY. Advanced glycation end products activate Smad signaling via TGF-beta-dependent and independent mechanisms: implications for diabetic renal and vascular disease. FASEB J. 2004;18:176–178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
