

The basis for haplotype complexity in VCBPs, an immune-type receptor in amphioxus
- PMID: 20652563
- PMCID: PMC3707128
- DOI: 10.1007/s00251-010-0464-x
The basis for haplotype complexity in VCBPs, an immune-type receptor in amphioxus
Abstract
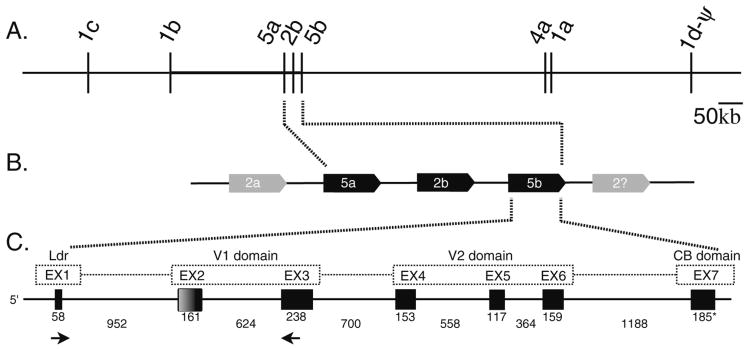
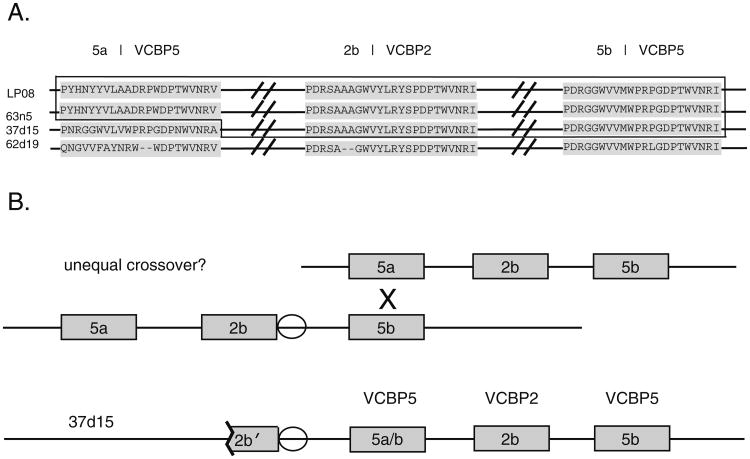
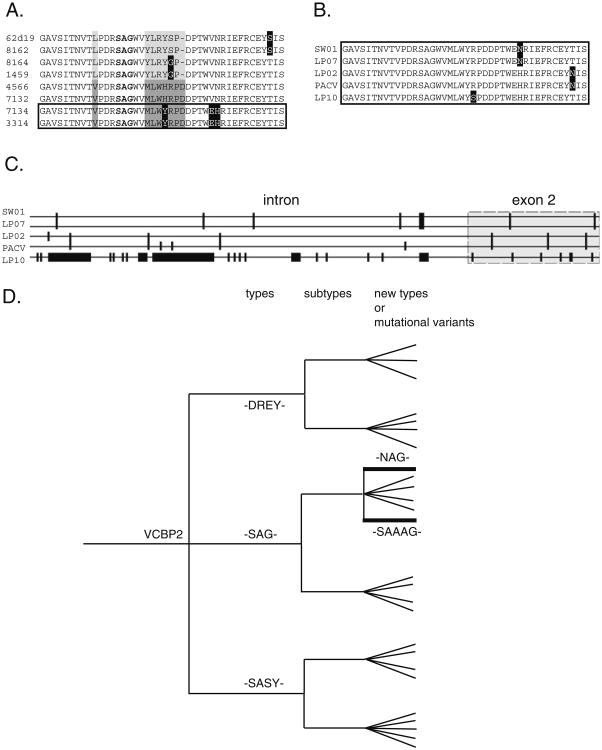
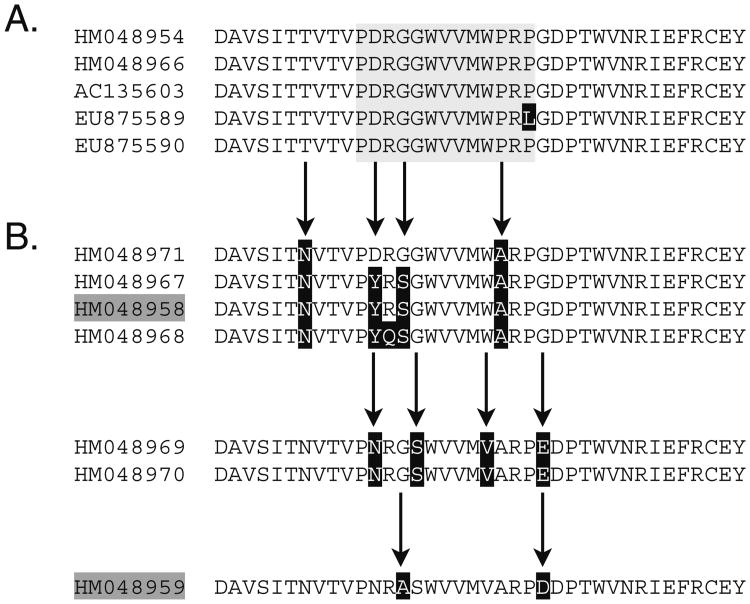
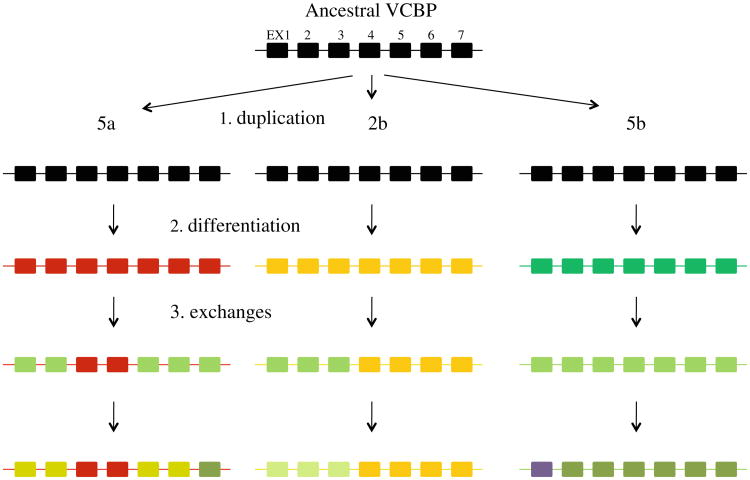
Innate immune gene repertoires are restricted primarily to germline variation. Adaptive immunity, by comparison, relies on somatic variation of germline-encoded genes to generate extraordinarily large numbers of non-heritable antigen recognition motifs. Invertebrates lack the key features of vertebrate adaptive immunity, but have evolved a variety of alternative mechanisms to successfully protect the integrity of "self"; in many cases, these appear to be taxon-specific innovations. In the protochordate Branchiostoma floridae (amphioxus), the variable region-containing chitin-binding proteins (VCBPs) constitute a multigene family (comprised of VCBPs 1-5), which possesses features that are consistent with innate immune-type function. A large number of VCBP alleles and haplotypes are shown to exhibit levels of polymorphism exceeding the elevated overall levels determined for the whole amphioxus genome (JGI). VCBP genes of the 2 and 5 types are distinguished further by a highly polymorphic segment (exon 2) in the N-terminal immunoglobulin domain, defined previously as a "hypervariable region" or a "hotspot." Genomic deoxyribonucleic acid (DNA) and complementary DNA (cDNA) sequences from large numbers of animals representing different populations reveal further significant differences in sequence complexity within and across VCBP2/5 haplotypes that arise through overlapping mechanisms of genetic exchange, gene copy number variation as well as mutation and give rise to distinct allelic lineages. The collective observations suggest that mechanisms were in place at the time of divergence of the cephalochordates that could selectively hyperdiversify immune-type receptors within a multigene family.
Figures
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Cannon JP, Haire RN, Litman GW. Identification of diversified genes that contain immunoglobulin-like variable regions in a protochordate. Nat Immunol. 2002;3:1200–1207. - PubMed
-
- Cannon JP, Haire RN, Rast JP, Litman GW. The phylogenetic origins of the antigen binding receptors and somatic diversification mechanisms. Immunol Rev. 2004a;200:12–22. - PubMed
-
- Cannon JP, Haire RN, Schnitker N, Mueller MG, Litman GW. Individual protochordates possess unique immune-type receptor repertoires. Curr Biol. 2004b;14:R465–R466. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
