

The M-domain controls Hsp104 protein remodeling activity in an Hsp70/Hsp40-dependent manner
- PMID: 20654624
- PMCID: PMC2938849
- DOI: 10.1016/j.jmb.2010.07.030
The M-domain controls Hsp104 protein remodeling activity in an Hsp70/Hsp40-dependent manner
Abstract
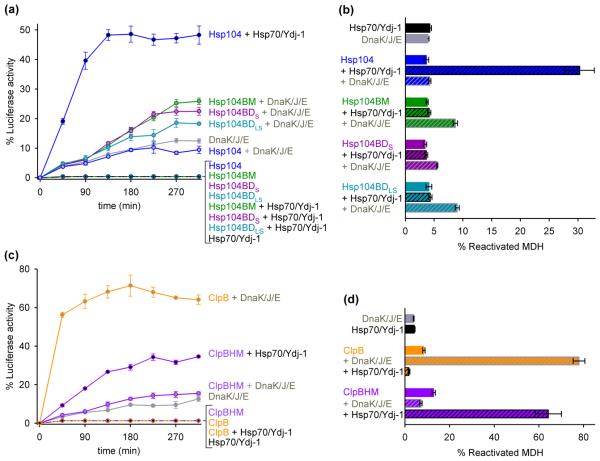
Yeast Hsp104 is a ring-forming ATP-dependent protein disaggregase that, together with the cognate Hsp70 chaperone system, has the remarkable ability to rescue stress-damaged proteins from a previously aggregated state. Both upstream and downstream functions for the Hsp70 system have been reported, but it remains unclear how Hsp70/Hsp40 is coupled to Hsp104 protein remodeling activity. Hsp104 is a multidomain protein that possesses an N-terminal domain, an M-domain, and two tandem AAA(+) domains. The M-domain forms an 85-A long coiled coil and is a hallmark of the Hsp104 chaperone family. While the three-dimensional structure of Hsp104 has been determined, the function of the M-domain is unclear. Here, we demonstrate that the M-domain is essential for protein disaggregation, but dispensable for Hsp104 ATPase- and substrate-translocating activities. Remarkably, replacing the Hsp104 M-domain with that of bacterial ClpB, and vice versa, switches species specificity so that our chimeras now cooperate with the noncognate Hsp70/DnaK chaperone system. Our results demonstrate that the M-domain controls Hsp104 protein remodeling activities in an Hsp70/Hsp40-dependent manner, which is required to unleash Hsp104 protein disaggregating activity.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures



Similar articles
-
Conserved distal loop residues in the Hsp104 and ClpB middle domain contact nucleotide-binding domain 2 and enable Hsp70-dependent protein disaggregation.J Biol Chem. 2014 Jan 10;289(2):848-67. doi: 10.1074/jbc.M113.520759. Epub 2013 Nov 26. J Biol Chem. 2014. PMID: 24280225 Free PMC article.
-
Structural determinants for protein unfolding and translocation by the Hsp104 protein disaggregase.Biosci Rep. 2017 Dec 22;37(6):BSR20171399. doi: 10.1042/BSR20171399. Print 2017 Dec 22. Biosci Rep. 2017. PMID: 29175998 Free PMC article.
-
Species-specific collaboration of heat shock proteins (Hsp) 70 and 100 in thermotolerance and protein disaggregation.Proc Natl Acad Sci U S A. 2011 Apr 26;108(17):6915-20. doi: 10.1073/pnas.1102828108. Epub 2011 Apr 7. Proc Natl Acad Sci U S A. 2011. PMID: 21474779 Free PMC article.
-
Chaperone networks in protein disaggregation and prion propagation.J Struct Biol. 2012 Aug;179(2):152-60. doi: 10.1016/j.jsb.2012.05.002. Epub 2012 May 10. J Struct Biol. 2012. PMID: 22580344 Review.
-
Chaperone-assisted protein aggregate reactivation: Different solutions for the same problem.Arch Biochem Biophys. 2015 Aug 15;580:121-34. doi: 10.1016/j.abb.2015.07.006. Epub 2015 Jul 6. Arch Biochem Biophys. 2015. PMID: 26159839 Review.
Cited by
-
Heat shock protein (Hsp) 70 is an activator of the Hsp104 motor.Proc Natl Acad Sci U S A. 2013 May 21;110(21):8513-8. doi: 10.1073/pnas.1217988110. Epub 2013 May 6. Proc Natl Acad Sci U S A. 2013. PMID: 23650362 Free PMC article.
-
Interaction of substrate-mimicking peptides with the AAA+ ATPase ClpB from Escherichia coli.Arch Biochem Biophys. 2018 Oct 1;655:12-17. doi: 10.1016/j.abb.2018.08.002. Epub 2018 Aug 6. Arch Biochem Biophys. 2018. PMID: 30092228 Free PMC article.
-
Hsp104 N-terminal domain interaction with substrates plays a regulatory role in protein disaggregation.FEBS J. 2022 Sep;289(17):5359-5377. doi: 10.1111/febs.16441. Epub 2022 Mar 30. FEBS J. 2022. PMID: 35305079 Free PMC article.
-
Comparative Analysis of the Structure and Function of AAA+ Motors ClpA, ClpB, and Hsp104: Common Threads and Disparate Functions.Front Mol Biosci. 2017 Aug 3;4:54. doi: 10.3389/fmolb.2017.00054. eCollection 2017. Front Mol Biosci. 2017. PMID: 28824920 Free PMC article. Review.
-
Hsp70 targets Hsp100 chaperones to substrates for protein disaggregation and prion fragmentation.J Cell Biol. 2012 Aug 6;198(3):387-404. doi: 10.1083/jcb.201201074. J Cell Biol. 2012. PMID: 22869599 Free PMC article.
References
-
- Bosl B, Grimminger V, Walter S. The molecular chaperone Hsp104--a molecular machine for protein disaggregation. J. Struct. Biol. 2006;156:139–148. - PubMed
-
- Doyle SM, Wickner S. Hsp104 and ClpB: protein disaggregating machines. Trends Biochem. Sci. 2009;34:40–48. - PubMed
-
- Haslberger T, Bukau B, Mogk A. Towards a unifying mechanism for ClpB/Hsp104-mediated protein disaggregation and prion propagation. Biochem. Cell Biol. 2010;88:63–75. - PubMed
-
- Chernoff YO, Lindquist SL, Ono B, Inge-Vechtomov SG, Liebman SW. Role of the chaperone protein Hsp104 in propagation of the yeast prion-like factor [PSI+] Science. 1995;268:880–884. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
