

The Bermuda Triangle of cocaine-induced neuroadaptations
- PMID: 20655604
- PMCID: PMC2935206
- DOI: 10.1016/j.tins.2010.06.003
The Bermuda Triangle of cocaine-induced neuroadaptations
Abstract
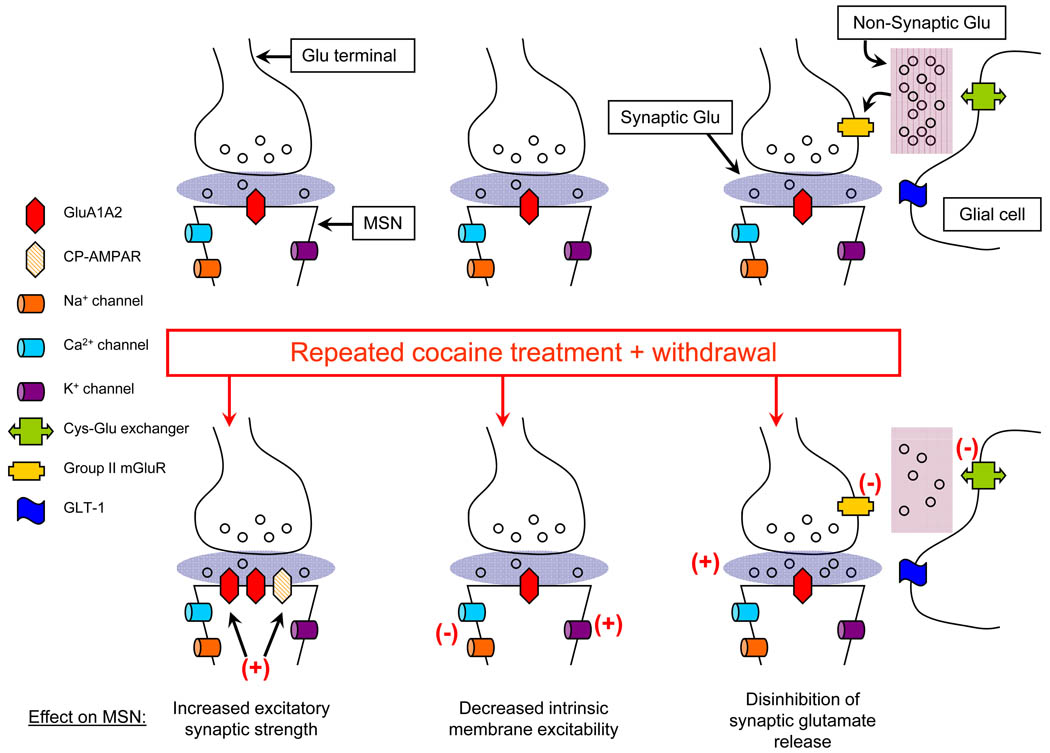
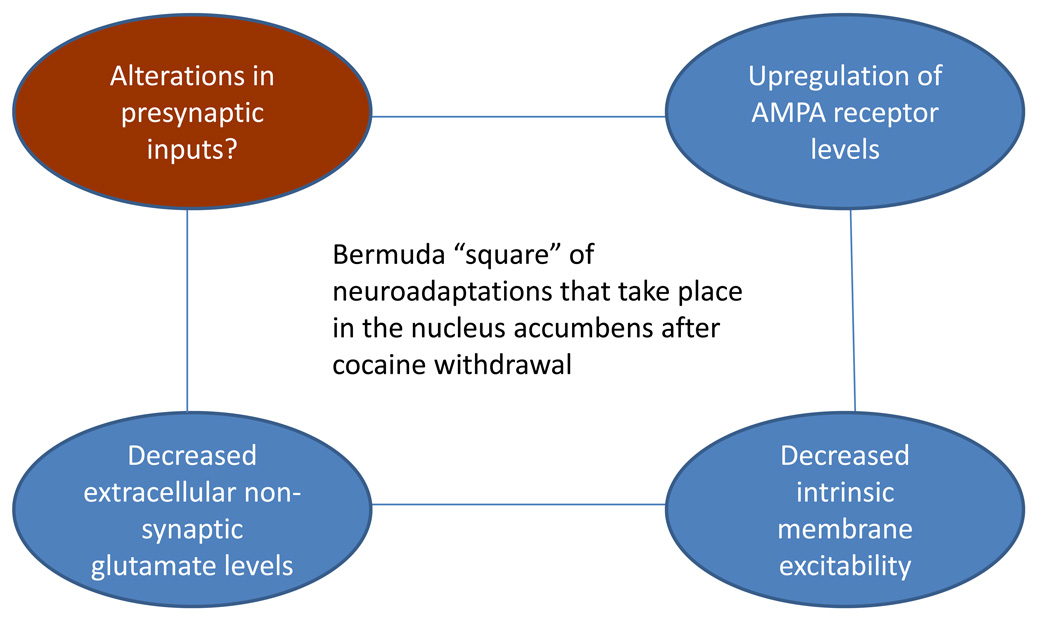
Activation of medium spiny neurons (MSNs) of the nucleus accumbens is critical for goal-directed behaviors including cocaine seeking. Studies in cocaine-experienced rodents have revealed three major categories of neuroadaptations that influence the ability of glutamate inputs to activate MSNs: changes in synaptic AMPA receptor levels, changes in extracellular non-synaptic glutamate levels and changes in MSN intrinsic membrane excitability. Most studies have focused on one of these adaptations. This review will consider the possibility that they are causally related and speculate about how time-dependent changes in their interactions may regulate MSN output during early and late withdrawal from repeated cocaine exposure.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Groenewegen HJ, et al. Convergence and segregation of ventral striatal inputs and outputs. Ann. N. Y. Acad. Sci. 1999;877:49–63. - PubMed
-
- Kelley AE. Functional specificity of ventral striatal compartments in appetitive behaviors. Ann. N.Y. Acad. Sci. 1999;877:71–90. - PubMed
-
- Kalivas PW, Volkow ND. The neural basis of addiction: a pathology of motivation and choice. Am. J. Psychiatry. 2005;162:1403–1413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
