

The potential landscape of genetic circuits imposes the arrow of time in stem cell differentiation
- PMID: 20655830
- PMCID: PMC2895388
- DOI: 10.1016/j.bpj.2010.03.058
The potential landscape of genetic circuits imposes the arrow of time in stem cell differentiation
Abstract
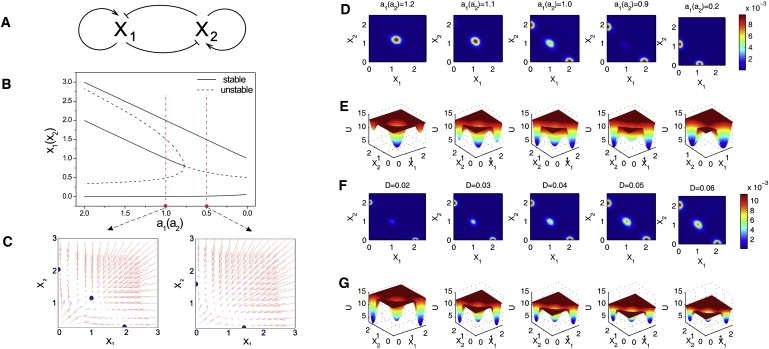
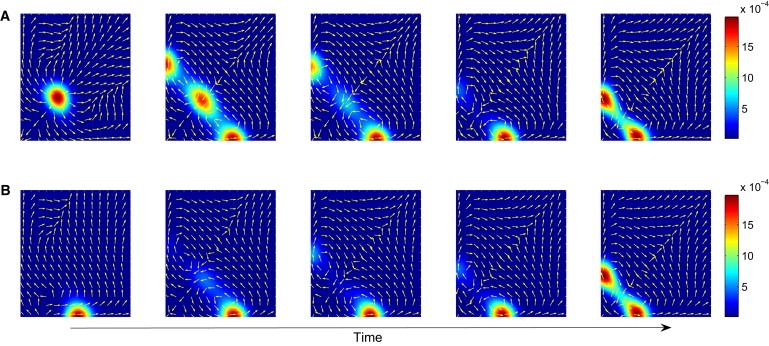
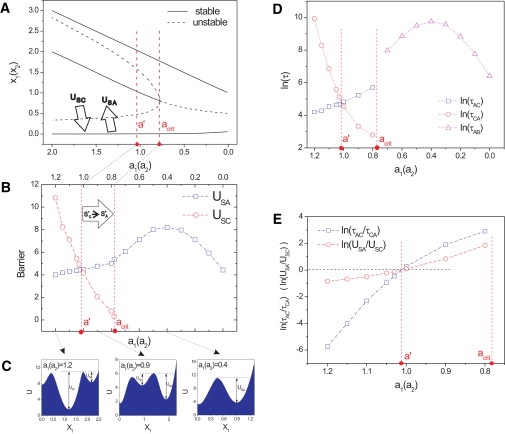
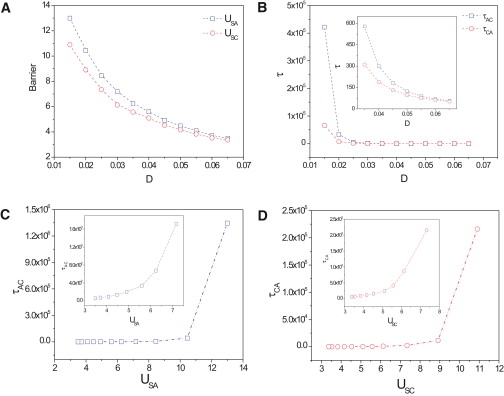
Differentiation from a multipotent stem or progenitor state to a mature cell is an essentially irreversible process. The associated changes in gene expression patterns exhibit time-directionality. This "arrow of time" in the collective change of gene expression across multiple stable gene expression patterns (attractors) is not explained by the regulated activation, the suppression of individual genes which are bidirectional molecular processes, or by the standard dynamical models of the underlying gene circuit which only account for local stability of attractors. To capture the global dynamics of this nonequilibrium system and gain insight in the time-asymmetry of state transitions, we computed the quasipotential landscape of the stochastic dynamics of a canonical gene circuit that governs branching cell fate commitment. The potential landscape reveals the global dynamics and permits the calculation of potential barriers between cell phenotypes imposed by the circuit architecture. The generic asymmetry of barrier heights indicates that the transition from the uncommitted multipotent state to differentiated states is inherently unidirectional. The model agrees with observations and predicts the extreme conditions for reprogramming cells back to the undifferentiated state.
Copyright 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Akashi K., He X., Li L. Transcriptional accessibility for genes of multiple tissues and hematopoietic lineages is hierarchically controlled during early hematopoiesis. Blood. 2003;101:383–389. - PubMed
-
- Huang S. Endothelial Biomedicine. Cambridge University Press; New York: 2007. Cell fates as attractors—stability and flexibility of cellular phenotype.
-
- Smith L., Greenfield A. DNA microarrays and development. Hum. Mol. Genet. 2003;12:R1–R8. - PubMed
-
- Kloc M., Zagrodzinska B. Chromatin elimination—an oddity or a common mechanism in differentiation and development? Differentiation. 2001;68:84–91. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
