

Actomyosin tension exerted on the nucleus through nesprin-1 connections influences endothelial cell adhesion, migration, and cyclic strain-induced reorientation
- PMID: 20655839
- PMCID: PMC2895377
- DOI: 10.1016/j.bpj.2010.04.011
Actomyosin tension exerted on the nucleus through nesprin-1 connections influences endothelial cell adhesion, migration, and cyclic strain-induced reorientation
Erratum in
- Biophys J. 2010 Aug 9;99(4):1329
Abstract
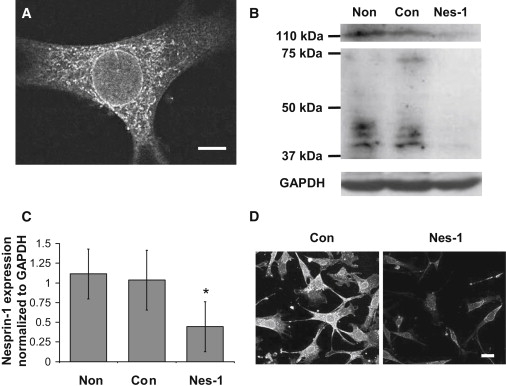
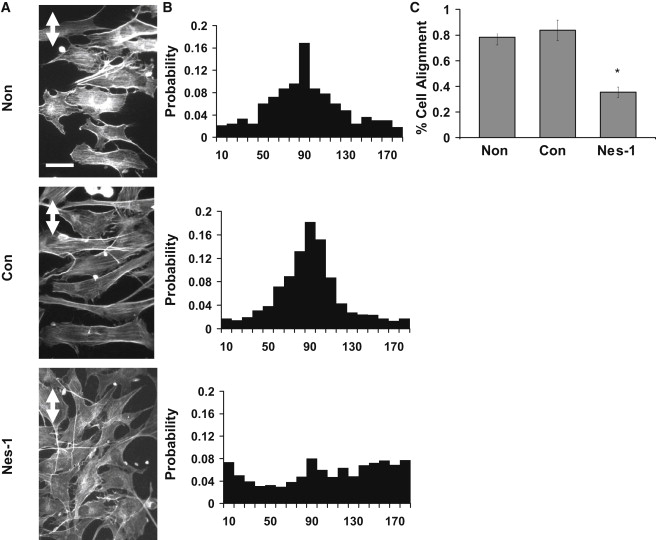
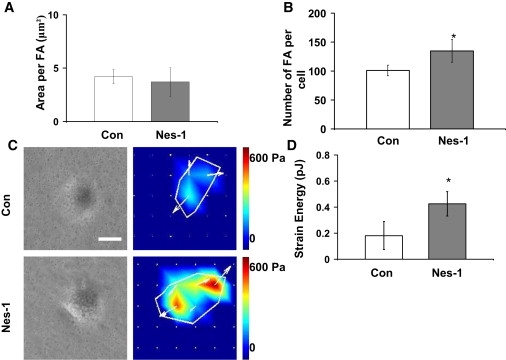
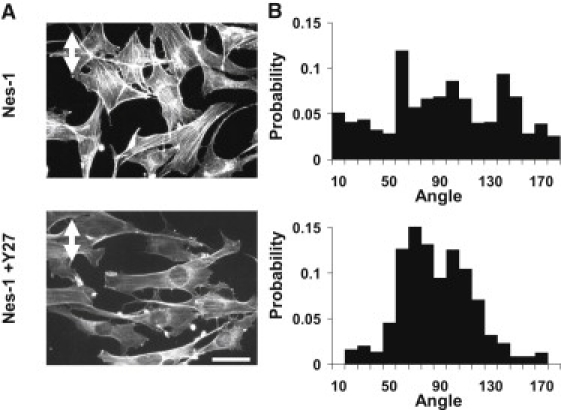
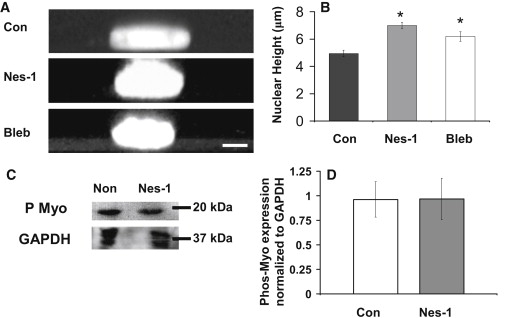
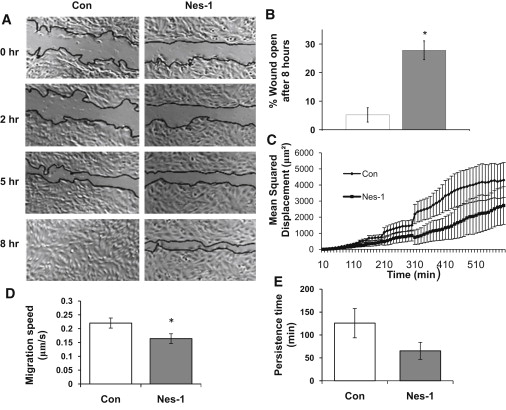
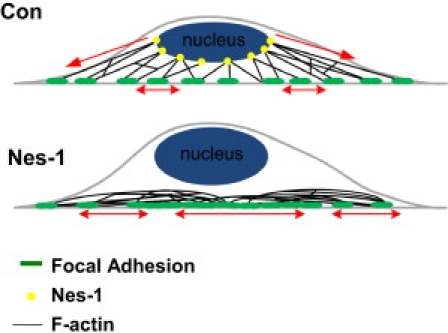
Endothelial cell polarization and directional migration is required for angiogenesis. Polarization and motility requires not only local cytoskeletal remodeling but also the motion of intracellular organelles such as the nucleus. However, the physiological significance of nuclear positioning in the endothelial cell has remained largely unexplored. Here, we show that siRNA knockdown of nesprin-1, a protein present in the linker of nucleus to cytoskeleton complex, abolished the reorientation of endothelial cells in response to cyclic strain. Confocal imaging revealed that the nuclear height is substantially increased in nesprin-1 depleted cells, similar to myosin inhibited cells. Nesprin-1 depletion increased the number of focal adhesions and substrate traction while decreasing the speed of cell migration; however, there was no detectable change in nonmuscle myosin II activity in nesprin-1 deficient cells. Together, these results are consistent with a model in which the nucleus balances a portion of the actomyosin tension in the cell. In the absence of nesprin-1, actomyosin tension is balanced by the substrate, leading to abnormal adhesion, migration, and cyclic strain-induced reorientation.
Copyright 2010 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Huang S., Ingber D.E. The structural and mechanical complexity of cell-growth control. Nat. Cell Biol. 1999;1:E131–E138. - PubMed
-
- Lauffenburger D.A., Horwitz A.F. Cell migration: a physically integrated molecular process. Cell. 1996;84:359–369. - PubMed
-
- Ferrara N., Gerber H.P., LeCouter J. The biology of VEGF and its receptors. Nat. Med. 2003;9:669–676. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
