

Centriole reduplication during prolonged interphase requires procentriole maturation governed by Plk1
- PMID: 20656208
- PMCID: PMC2911792
- DOI: 10.1016/j.cub.2010.05.050
Centriole reduplication during prolonged interphase requires procentriole maturation governed by Plk1
Abstract
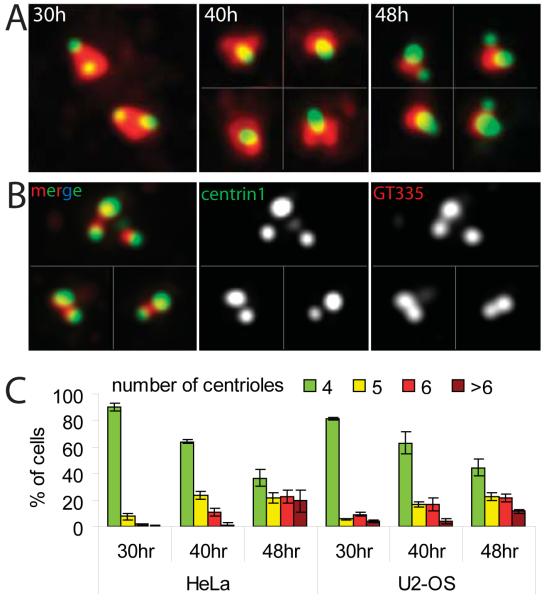
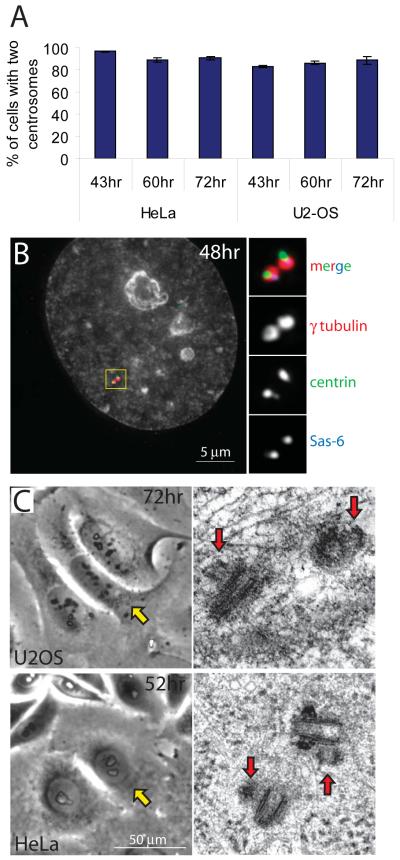
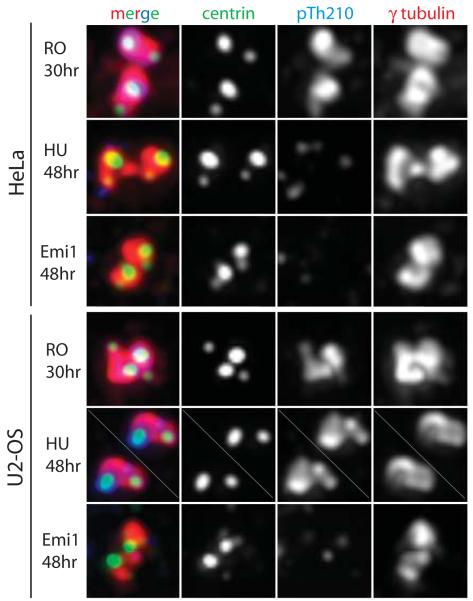
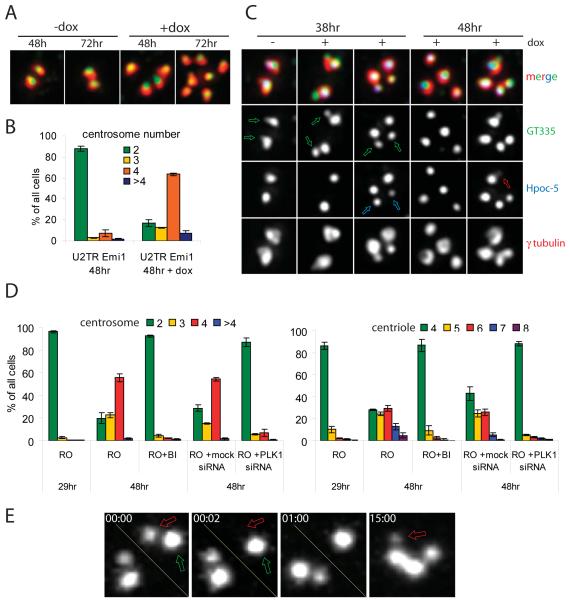
Supernumerary centrioles lead to abnormal mitosis, which in turn promotes tumorigenesis. Thus, centriole duplication must be coordinated with the cell cycle to ensure that the number of centrioles in the cell doubles precisely during each cell cycle. However, in some transformed cells, centrioles undergo multiple rounds of duplication (reduplication) during prolonged interphase. Mechanisms responsible for centriole reduplication are poorly understood. Here, we report that centrioles reduplicate consistently in cancerous and nontransformed human cells during G2 arrests and that this reduplication requires the activity of Polo-like kinase 1 (Plk1). We also find that a cell's ability to reduplicate centrioles during S arrests depends on the presence of activated (Thr210-phosphorylated) Plk1 at the centrosome. In the absence of activated Plk1, nascent procentrioles remain associated with mother centrioles, which prevents centriole reduplication. In contrast, if Plk1(pT210) appears at the centrosome, procentrioles mature, disengage from mother centrioles, and ultimately duplicate. Plk1 activity is not required for the assembly of procentrioles, however. Thus, the role of Plk1 is to coordinate the centriole duplication cycle with the cell cycle. Activation of Plk1 during late S/G2 induces procentriole maturation, and after this point, the centriole cycle can be completed autonomously, even in the absence of cell-cycle progression.
2010 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
