

Chitin particles are multifaceted immune adjuvants
- PMID: 20656945
- PMCID: PMC3029935
- DOI: 10.1164/rccm.200912-1877OC
Chitin particles are multifaceted immune adjuvants
Abstract
Rationale: Chitin is a ubiquitous polysaccharide in fungi, insects, allergens, and parasites that is released at sites of infection. Its role in the generation of tissue inflammation, however, is not fully understood.
Objectives: We hypothesized that chitin is an important adjuvant for adaptive immunity.
Methods: Mice were injected with a solution of ovalbumin and chitin.
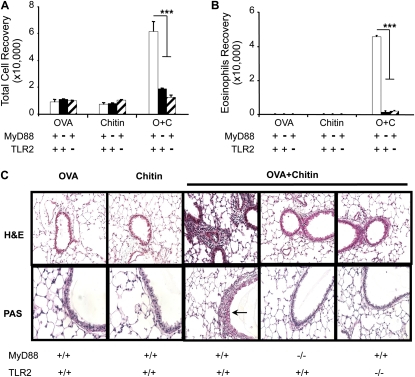
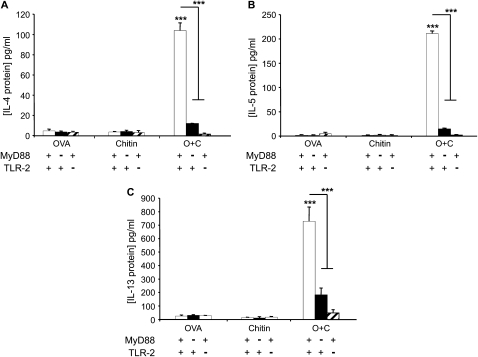
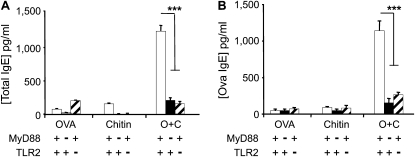
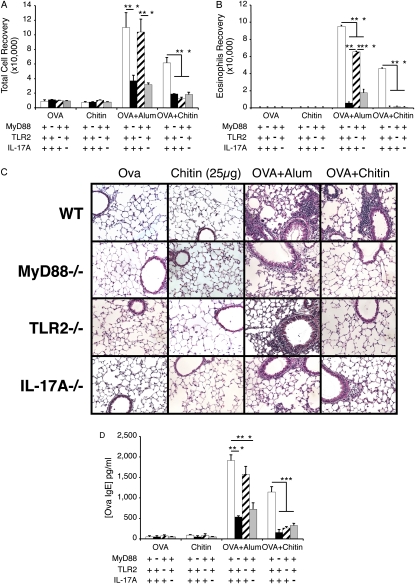
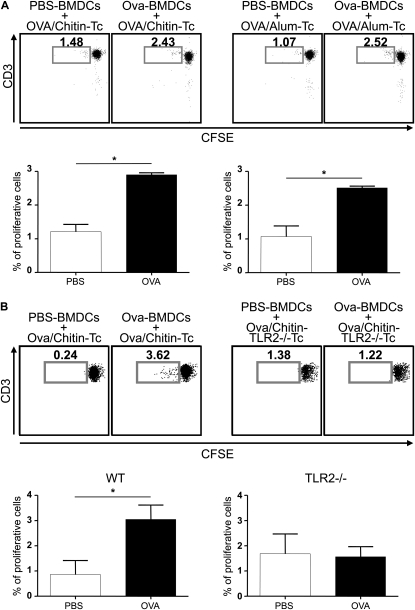
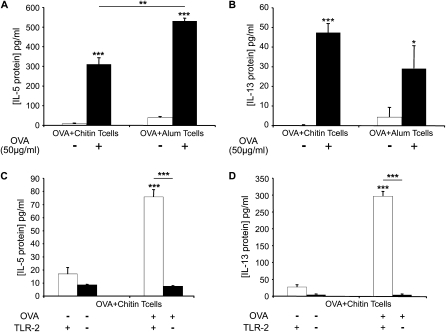
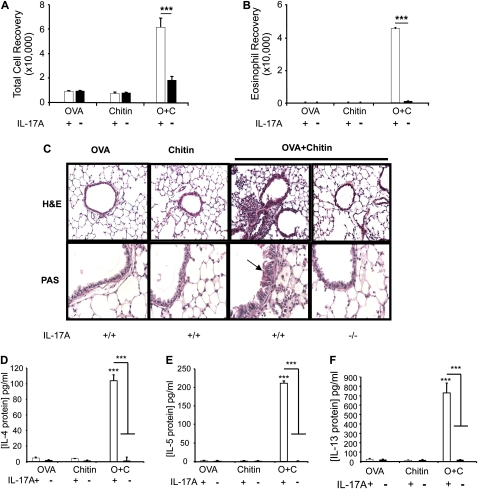
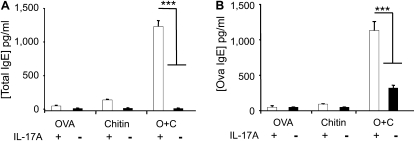
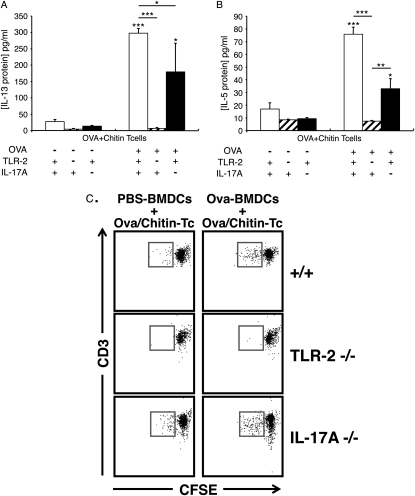
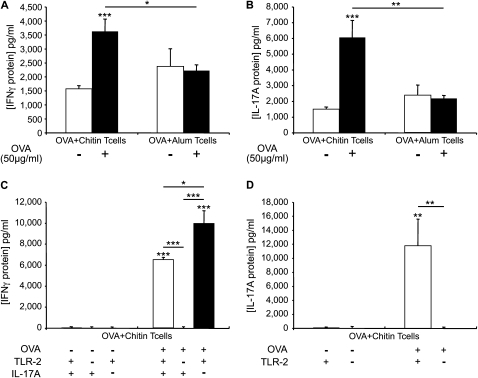
Measurements and main results: We used in vivo and ex vivo/in vitro approaches to characterize the ability of chitin fragments to foster adaptive immune responses against ovalbumin and compared these responses to those induced by aluminum hydroxide (alum). In vivo, ovalbumin challenge caused an eosinophil-rich pulmonary inflammatory response, Th2 cytokine elaboration, IgE induction, and mucus metaplasia in mice that had been sensitized with ovalbumin plus chitin or ovalbumin plus alum. Toll-like receptor-2, MyD88, and IL-17A played critical roles in the chitin-induced responses, and MyD88 and IL-17A played critical roles in the alum-induced responses. In vitro, CD4(+) T cells from mice sensitized with ovalbumin plus chitin were incubated with ovalbumin-stimulated bone marrow-derived dendritic cells. In these experiments, CD4(+) T-cell proliferation, IL-5, IL-13, IFN-γ, and IL-17A production were appreciated. Toll-like receptor-2, MyD88, and IL-17A played critical roles in these in vitro adjuvant properties of chitin. TLR-2 was required for cell proliferation, whereas IL-17 and TLR-2 were required for cytokine elaboration. IL-17A also inhibited the generation of adaptive Th1 responses.
Conclusions: These studies demonstrate that chitin is a potent multifaceted adjuvant that induces adaptive Th2, Th1, and Th17 immune responses. They also demonstrate that the adjuvant properties of chitin are mediated by a pathway(s) that involves and is regulated by TLR-2, MyD88, and IL-17A.
Figures
References
-
- Neville AC, Parry DA, Woodhead-Galloway J. The chitin crystallite in arthropod cuticle. J Cell Sci 1976;21:73. - PubMed
-
- Fuhrman JA, Piessens WF. Chitin synthesis and sheath morphogenesis in Brugia malayi microfilariae. Mol Biochem Parasitol 1985;17:93. - PubMed
-
- Araujo AC, Souto-Padron T, de Souza W. Cytochemical localization of carbohydrate residues in microfilariae of Wuchereria bancrofti and Brugia malayi. J Histochem Cytochem 1993;41:571. - PubMed
-
- Debono M, Gordee RS. Antibiotics that inhibit fungal cell wall development. Annu Rev Microbiol 1994;48:471. - PubMed
-
- Zhu Z, Zheng T, Homer RJ, Kim YK, Chen NY, Cohn L, Hamid Q, Elias JA. Acidic mammalian chitinase in asthmatic Th2 inflammation and IL-13 pathway activation. Science 2004;304:1678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
