

Do humans optimally exploit redundancy to control step variability in walking?
- PMID: 20657664
- PMCID: PMC2904769
- DOI: 10.1371/journal.pcbi.1000856
Do humans optimally exploit redundancy to control step variability in walking?
Abstract
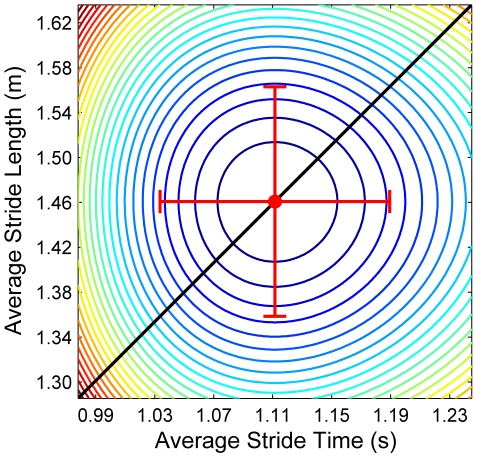
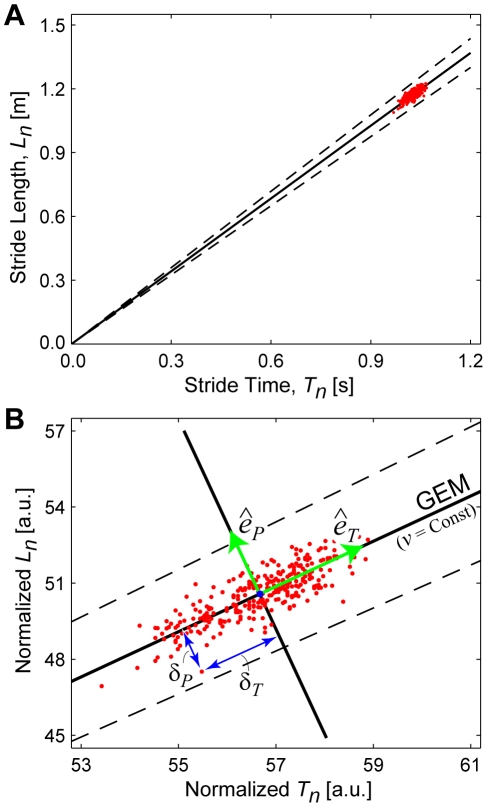
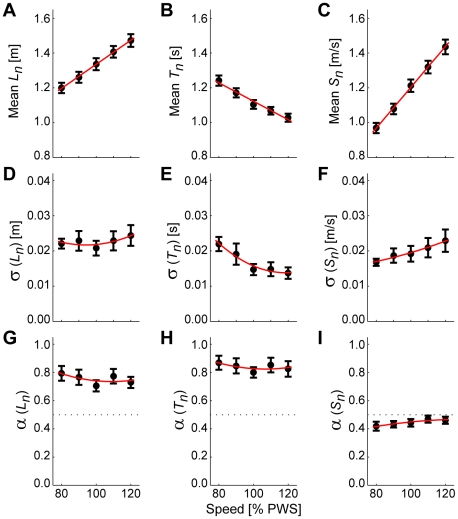
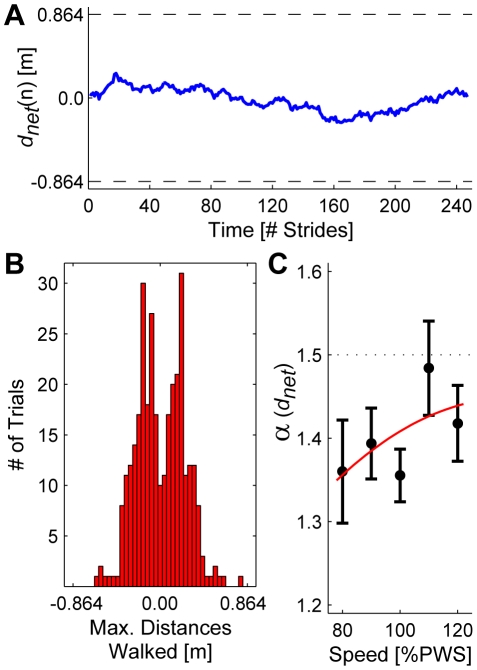
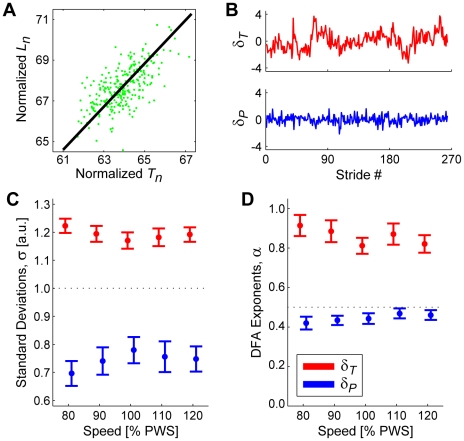
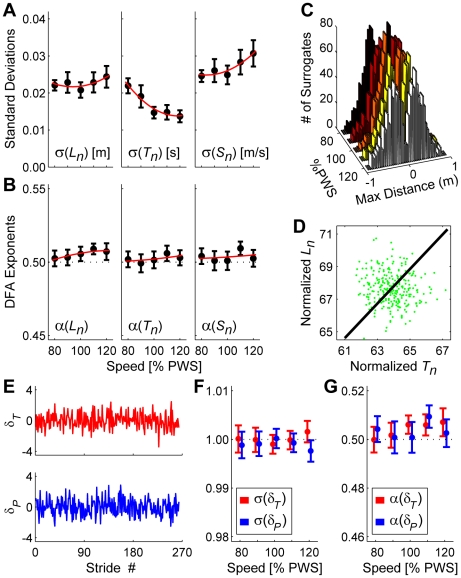
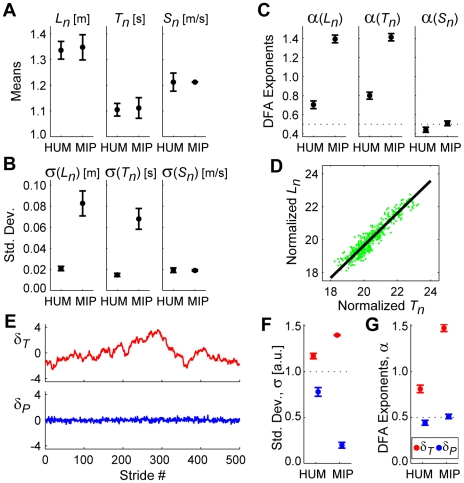
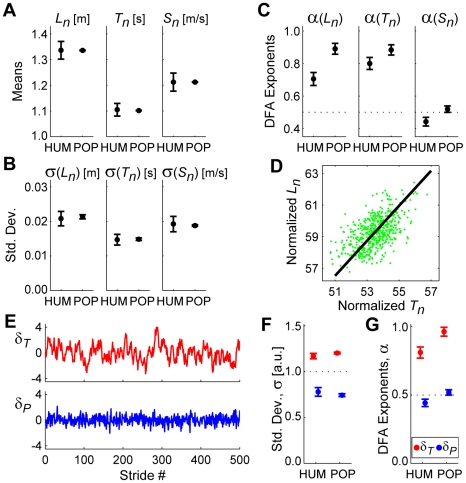
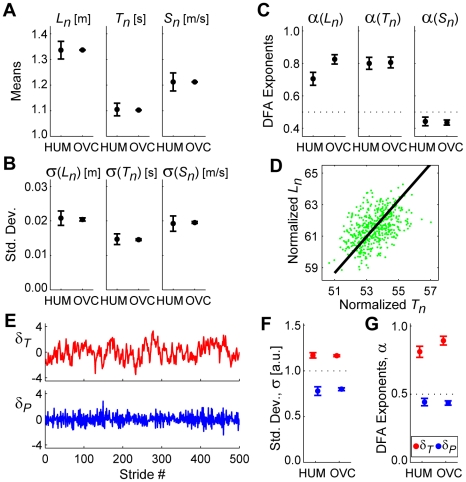
It is widely accepted that humans and animals minimize energetic cost while walking. While such principles predict average behavior, they do not explain the variability observed in walking. For robust performance, walking movements must adapt at each step, not just on average. Here, we propose an analytical framework that reconciles issues of optimality, redundancy, and stochasticity. For human treadmill walking, we defined a goal function to formulate a precise mathematical definition of one possible control strategy: maintain constant speed at each stride. We recorded stride times and stride lengths from healthy subjects walking at five speeds. The specified goal function yielded a decomposition of stride-to-stride variations into new gait variables explicitly related to achieving the hypothesized strategy. Subjects exhibited greatly decreased variability for goal-relevant gait fluctuations directly related to achieving this strategy, but far greater variability for goal-irrelevant fluctuations. More importantly, humans immediately corrected goal-relevant deviations at each successive stride, while allowing goal-irrelevant deviations to persist across multiple strides. To demonstrate that this was not the only strategy people could have used to successfully accomplish the task, we created three surrogate data sets. Each tested a specific alternative hypothesis that subjects used a different strategy that made no reference to the hypothesized goal function. Humans did not adopt any of these viable alternative strategies. Finally, we developed a sequence of stochastic control models of stride-to-stride variability for walking, based on the Minimum Intervention Principle. We demonstrate that healthy humans are not precisely "optimal," but instead consistently slightly over-correct small deviations in walking speed at each stride. Our results reveal a new governing principle for regulating stride-to-stride fluctuations in human walking that acts independently of, but in parallel with, minimizing energetic cost. Thus, humans exploit task redundancies to achieve robust control while minimizing effort and allowing potentially beneficial motor variability.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Zehr EP, Stein RB. What Functions do Reflexes Serve During Human Locomotion? Prog Neurobiol. 1999;58:185–205. - PubMed
-
- Warren WH, Kay BA, Zosh WD, Duchon AP, Sahuc S. Optic Flow is Used to Control Human Walking. Nat Neurosci. 2001;4:213–216. - PubMed
-
- Bent LR, Inglis JT, McFadyen BJ. When is Vestibular Information Important During Walking? J Neurophysiol. 2004;92:1269–1275. - PubMed
-
- Rossignol S, Dubuc R, Gossard J-P. Dynamic Sensorimotor Interactions in Locomotion. Physiol Rev. 2006;86:89–154. - PubMed
-
- Margaria R. Sulla fisiologia, e specialmente sul consumo energetico, della marcia e della corsa a varie velocita ed inclinazioni del terreno. Accad Naz Lincei Rc. 1938;6 7:299–368.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
