

A eubacterial riboswitch class that senses the coenzyme tetrahydrofolate
- PMID: 20659680
- PMCID: PMC3417113
- DOI: 10.1016/j.chembiol.2010.05.020
A eubacterial riboswitch class that senses the coenzyme tetrahydrofolate
Abstract
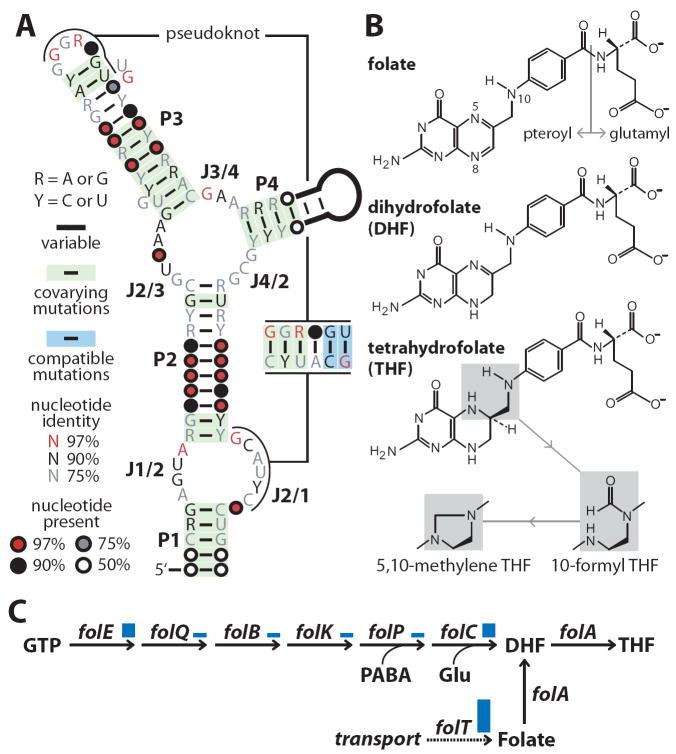
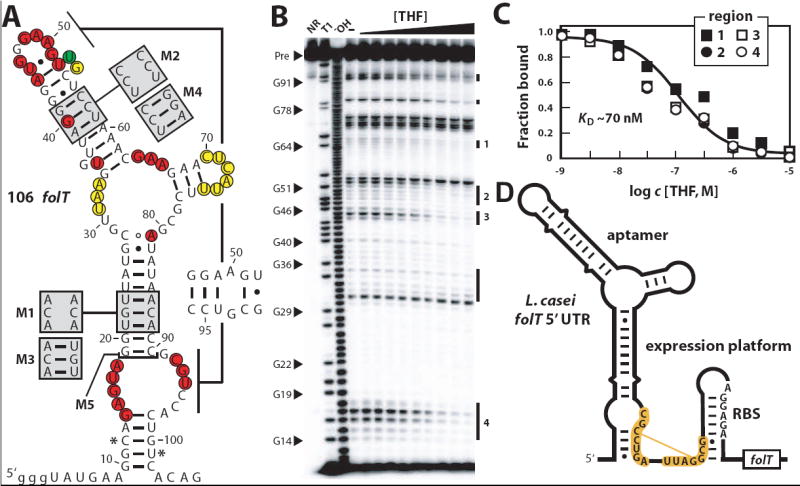
Comparative sequence analyses of bacterial genomes are revealing many structured RNA motifs that function as metabolite-binding riboswitches. We have identified an RNA motif frequently positioned in the 5' UTRs of folate transport and biosynthesis genes in Firmicute genomes. Biochemical experiments confirm that representatives of this new-found RNA class selectively bind derivatives of the vitamin folate, including di- and tetrahydrofolate coenzymes. In addition, representatives of this aptamer class occasionally reside upstream of RNA structures that are predicted to control translation initiation in response to ligand binding. These findings expand the number of coenzymes that are directly sensed by RNA and reveal possible riboswitch-controlled regulons that respond to changes in single-carbon metabolism.
2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Breaker RR. Complex riboswitches. Science. 2008;319:1795–1797. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
