

Novel Zn2+-binding sites in human transthyretin: implications for amyloidogenesis and retinol-binding protein recognition
- PMID: 20659897
- PMCID: PMC2951245
- DOI: 10.1074/jbc.M110.157206
Novel Zn2+-binding sites in human transthyretin: implications for amyloidogenesis and retinol-binding protein recognition
Abstract
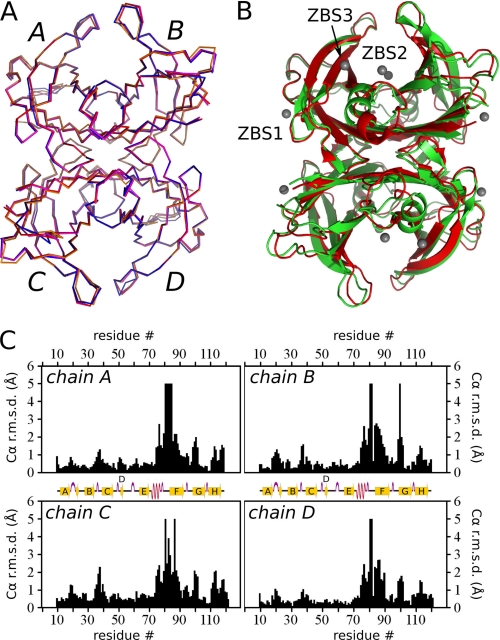
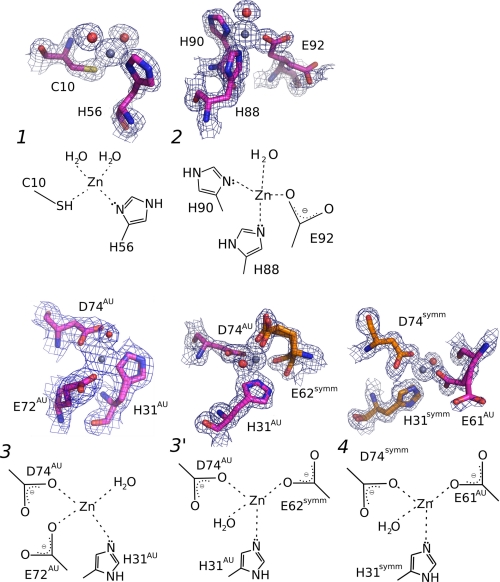
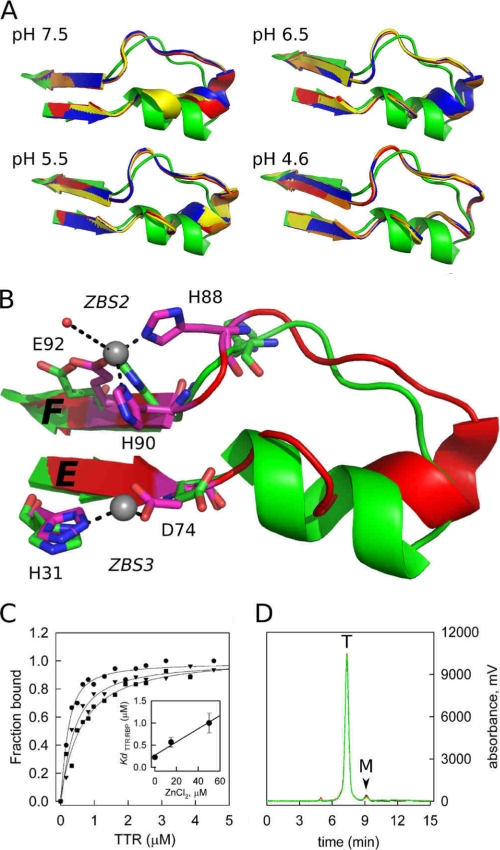
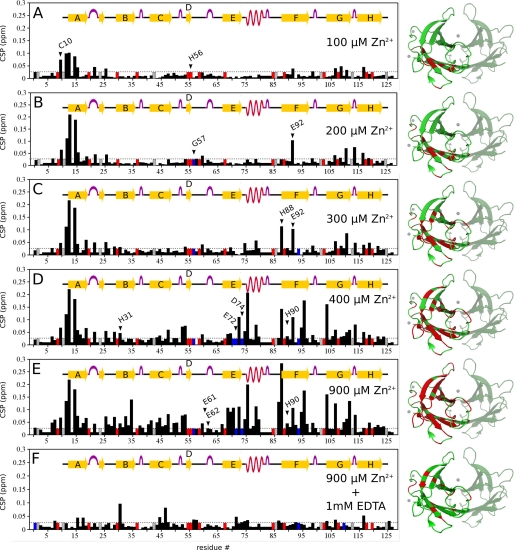
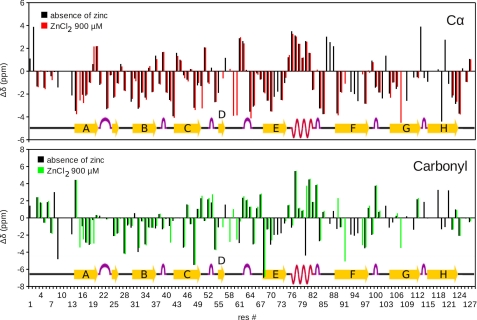
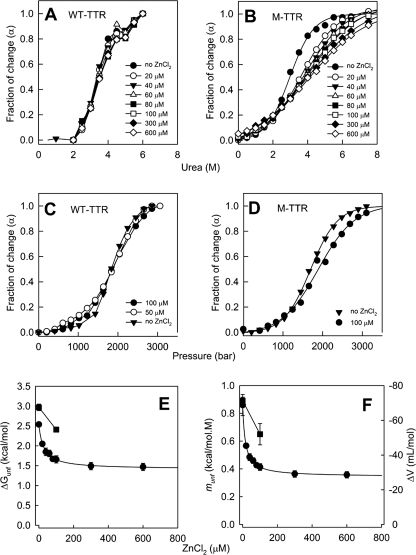
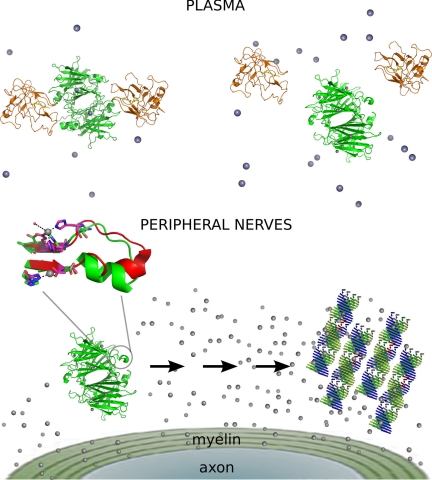
Human transthyretin (TTR) is a homotetrameric protein involved in several amyloidoses. Zn(2+) enhances TTR aggregation in vitro, and is a component of ex vivo TTR amyloid fibrils. We report the first crystal structure of human TTR in complex with Zn(2+) at pH 4.6-7.5. All four structures reveal three tetra-coordinated Zn(2+)-binding sites (ZBS 1-3) per monomer, plus a fourth site (ZBS 4) involving amino acid residues from a symmetry-related tetramer that is not visible in solution by NMR. Zn(2+) binding perturbs loop E-α-helix-loop F, the region involved in holo-retinol-binding protein (holo-RBP) recognition, mainly at acidic pH; TTR affinity for holo-RBP decreases ∼5-fold in the presence of Zn(2+). Interestingly, this same region is disrupted in the crystal structure of the amyloidogenic intermediate of TTR formed at acidic pH in the absence of Zn(2+). HNCO and HNCA experiments performed in solution at pH 7.5 revealed that upon Zn(2+) binding, although the α-helix persists, there are perturbations in the resonances of the residues that flank this region, suggesting an increase in structural flexibility. While stability of the monomer of TTR decreases in the presence of Zn(2+), which is consistent with the tertiary structural perturbation provoked by Zn(2+) binding, tetramer stability is only marginally affected by Zn(2+). These data highlight structural and functional roles of Zn(2+) in TTR-related amyloidoses, as well as in holo-RBP recognition and vitamin A homeostasis.
Figures
References
-
- Palha J. A. (2002) Clin. Chem. Lab. Med. 40, 1292–1300 - PubMed
-
- Monaco H. L. (2000) Biochim. Biophys. Acta 1482, 65–72 - PubMed
-
- Sekijima Y., Kelly J. W., Ikeda S. (2008) Curr. Pharm. Des. 14, 3219–3230 - PubMed
-
- Kingsbury J. S., Théberge R., Karbassi J. A., Lim A., Costello C. E., Connors L. H. (2007) Anal. Chem. 79, 1990–1998 - PubMed
-
- Inoue S., Kuroiwa M., Saraiva M. J., Guimarães A., Kisilevsky R. (1998) J. Struct. Biol. 124, 1–12 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
