

Ubc4 and Not4 regulate steady-state levels of DNA polymerase-α to promote efficient and accurate DNA replication
- PMID: 20660159
- PMCID: PMC2938386
- DOI: 10.1091/mbc.E09-06-0452
Ubc4 and Not4 regulate steady-state levels of DNA polymerase-α to promote efficient and accurate DNA replication
Abstract
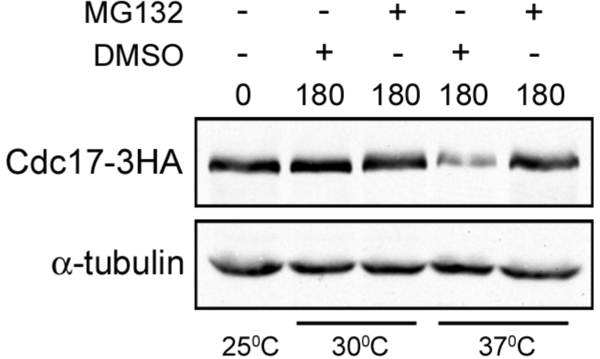
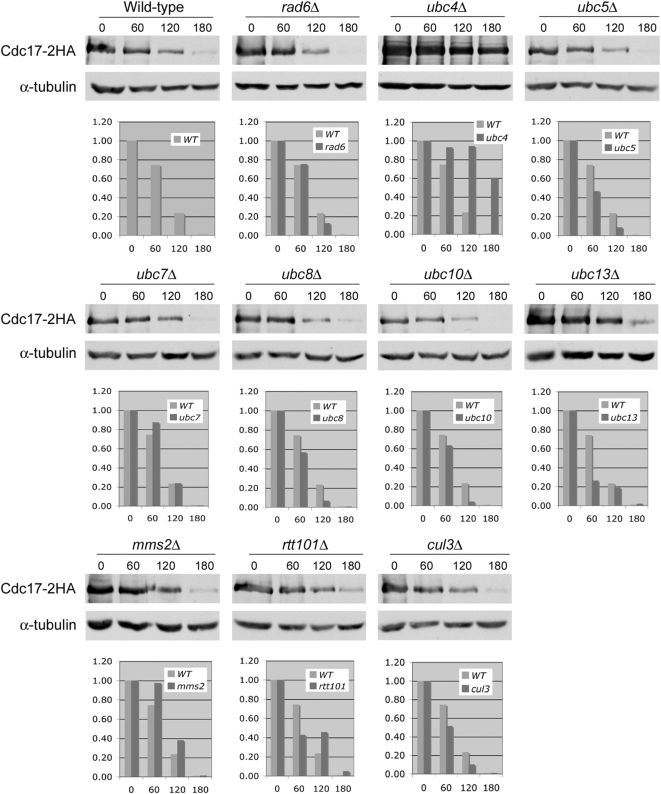
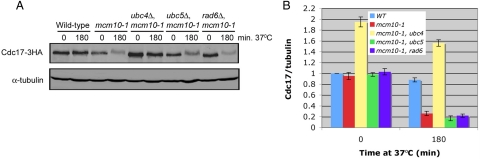
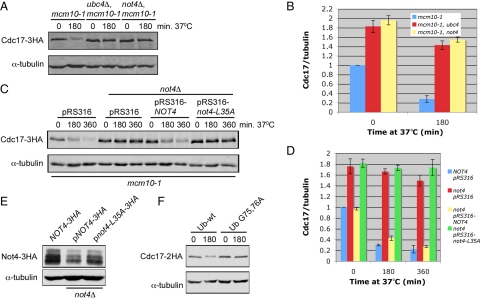
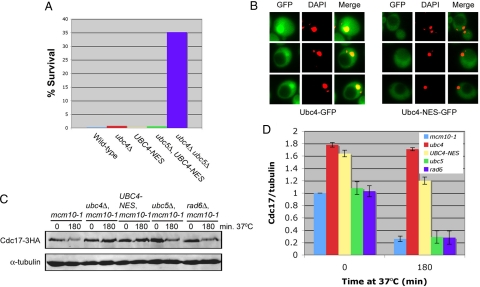
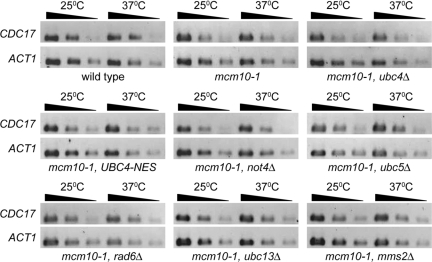
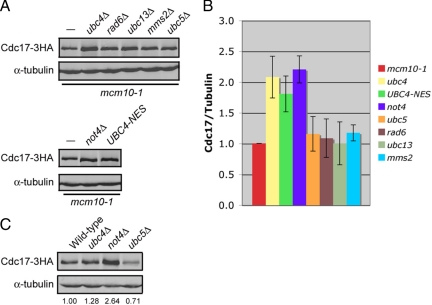
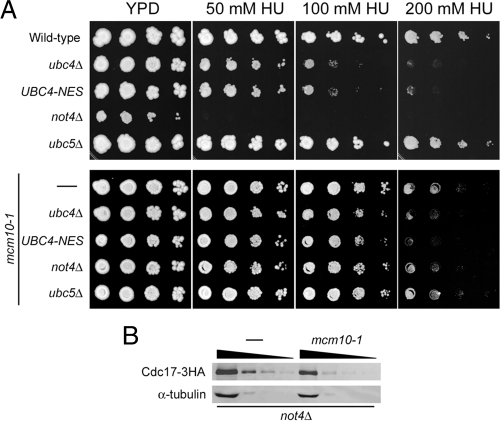
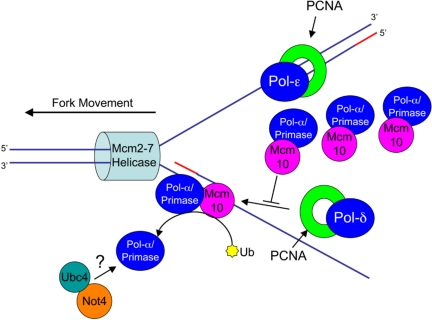
The accurate duplication of chromosomal DNA is required to maintain genomic integrity. However, from an evolutionary point of view, a low mutation rate during DNA replication is desirable. One way to strike the right balance between accuracy and limited mutagenesis is to use a DNA polymerase that lacks proofreading activity but contributes to DNA replication in a very restricted manner. DNA polymerase-α fits this purpose exactly, but little is known about its regulation at the replication fork. Minichromosome maintenance protein (Mcm) 10 regulates the stability of the catalytic subunit of pol-α in budding yeast and human cells. Cdc17, the catalytic subunit of pol-α in yeast, is rapidly degraded after depletion of Mcm10. Here we show that Ubc4 and Not4 are required for Cdc17 destabilization. Disruption of Cdc17 turnover resulted in sensitivity to hydroxyurea, suggesting that this pathway is important for DNA replication. Furthermore, overexpression of Cdc17 in ubc4 and not4 mutants caused slow growth and synthetic dosage lethality, respectively. Our data suggest that Cdc17 levels are very tightly regulated through the opposing forces of Ubc4 and Not4 (destabilization) and Mcm10 (stabilization). We conclude that regular turnover of Cdc17 via Ubc4 and Not4 is required for proper cell proliferation.
Figures
Similar articles
-
A conserved Hsp10-like domain in Mcm10 is required to stabilize the catalytic subunit of DNA polymerase-alpha in budding yeast.J Biol Chem. 2006 Jul 7;281(27):18414-25. doi: 10.1074/jbc.M513551200. Epub 2006 May 4. J Biol Chem. 2006. PMID: 16675460
-
Alternative mechanisms for coordinating polymerase alpha and MCM helicase.Mol Cell Biol. 2010 Jan;30(2):423-35. doi: 10.1128/MCB.01240-09. Epub 2009 Nov 16. Mol Cell Biol. 2010. PMID: 19917723 Free PMC article.
-
Mcm10 regulates the stability and chromatin association of DNA polymerase-alpha.Mol Cell. 2004 Oct 22;16(2):173-85. doi: 10.1016/j.molcel.2004.09.017. Mol Cell. 2004. PMID: 15494305
-
New Insights into the Mechanism of DNA Duplication by the Eukaryotic Replisome.Trends Biochem Sci. 2016 Oct;41(10):859-871. doi: 10.1016/j.tibs.2016.07.011. Epub 2016 Aug 20. Trends Biochem Sci. 2016. PMID: 27555051 Review.
-
Dynamic ubiquitin signaling in cell cycle regulation.J Cell Biol. 2017 Aug 7;216(8):2259-2271. doi: 10.1083/jcb.201703170. Epub 2017 Jul 6. J Cell Biol. 2017. PMID: 28684425 Free PMC article. Review.
Cited by
-
Mcm10 self-association is mediated by an N-terminal coiled-coil domain.PLoS One. 2013 Jul 23;8(7):e70518. doi: 10.1371/journal.pone.0070518. Print 2013. PLoS One. 2013. PMID: 23894664 Free PMC article.
-
The N-terminus of Mcm10 is important for interaction with the 9-1-1 clamp and in resistance to DNA damage.Nucleic Acids Res. 2014 Jul;42(13):8389-404. doi: 10.1093/nar/gku479. Epub 2014 Jun 27. Nucleic Acids Res. 2014. PMID: 24972833 Free PMC article.
-
MutSα mismatch repair protein stability is governed by subunit interaction, acetylation, and ubiquitination.G3 (Bethesda). 2021 Apr 23;11(3):jkaa065. doi: 10.1093/g3journal/jkaa065. G3 (Bethesda). 2021. PMID: 33793773 Free PMC article.
-
Ubiquitination-dependent control of sexual differentiation in fission yeast.Elife. 2017 Aug 25;6:e28046. doi: 10.7554/eLife.28046. Elife. 2017. PMID: 28841135 Free PMC article.
-
The C-terminal region of yeast ubiquitin-protein ligase Not4 mediates its cellular localization and stress response.FEMS Microbiol Lett. 2021 Aug 17;368(15):fnab097. doi: 10.1093/femsle/fnab097. FEMS Microbiol Lett. 2021. PMID: 34338747 Free PMC article.
References
-
- Aves S. J., Tongue N., Foster A. J., Hart E. A. The essential Schizosaccharomyces pombe cdc23 DNA replication gene shares structural and functional homology with the Saccharomyces cerevisiae DNA43 (MCM10) gene. Curr. Genetics. 1998;34:164–171. - PubMed
-
- Bell S. P., Dutta A. DNA replication in eukaryotic cells. Annual Rev. Biochem. 2002;71:333–374. - PubMed
-
- Bochman M. L., Schwacha A. The Mcm2–7 complex has in vitro helicase activity. Molecular Cell. 2008;31:287–293. - PubMed
-
- Boeke J. D., LaCroute F., Fink G. R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol. Gen. Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
