

The human basal ganglia modulate frontal-posterior connectivity during attention shifting
- PMID: 20660273
- PMCID: PMC6632831
- DOI: 10.1523/JNEUROSCI.1111-10.2010
The human basal ganglia modulate frontal-posterior connectivity during attention shifting
Abstract
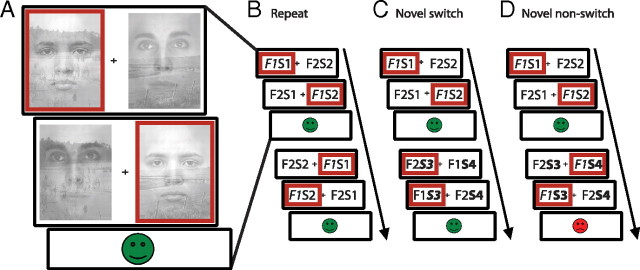
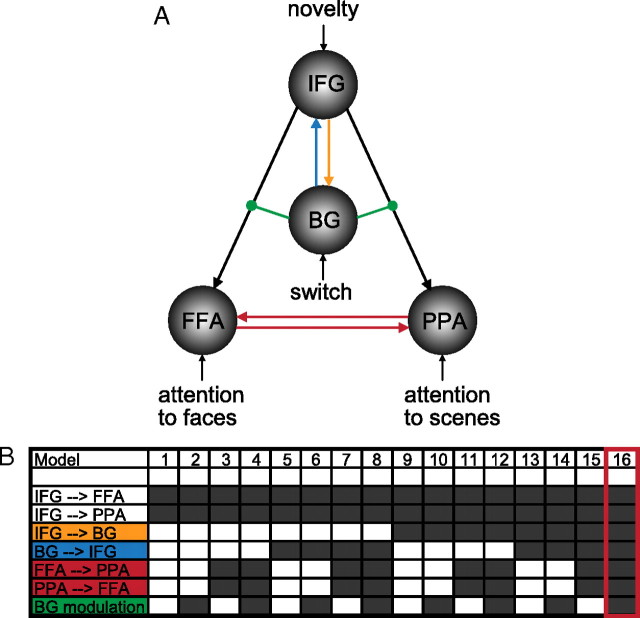
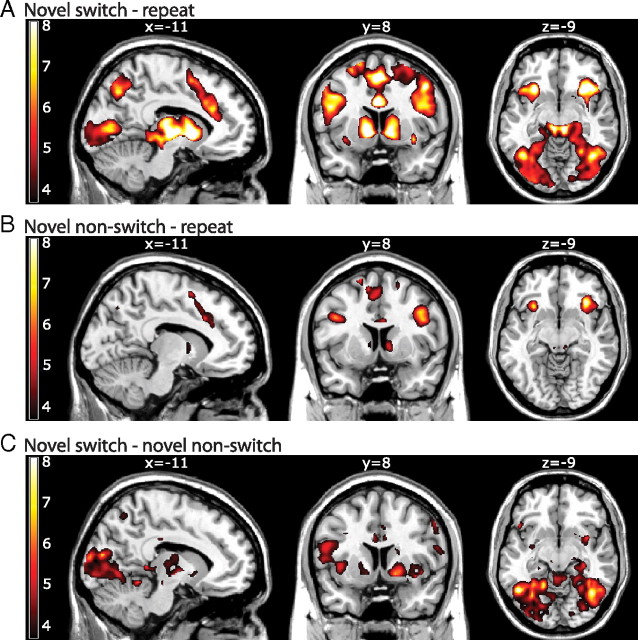
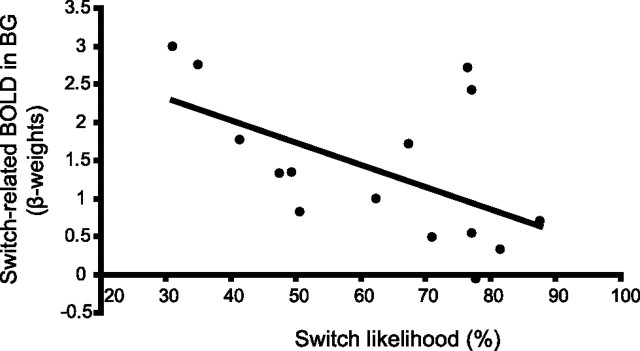
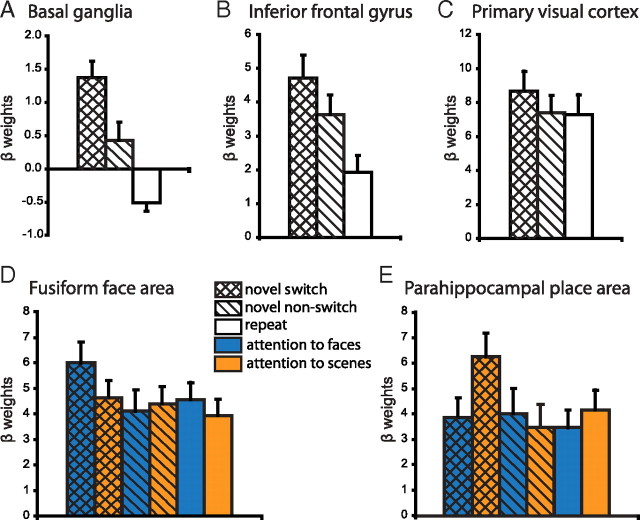
Current models of flexible cognitive control emphasize the role of the prefrontal cortex. This region has been shown to control attention by biasing information processing in favor of task-relevant representations. However, the prefrontal cortex does not act in isolation. We used functional magnetic resonance imaging combined with nonlinear dynamic causal modeling to demonstrate that the basal ganglia play a role in modulating the top-down influence of the prefrontal cortex on visual processing in humans. Specifically, our results reveal that connectivity between the prefrontal cortex and stimulus-specific visual association areas depends on activity in the ventral striatopallidum, elicited by salient events leading to shifts in attention. These data integrate disparate literatures on top-down control by the prefrontal cortex and selective gating by the basal ganglia and highlight the importance of the basal ganglia for high-level cognitive control.
Figures
References
-
- Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. 1986;9:357–381. - PubMed
-
- Braver TS, Cohen JD. On the control of control: the role of dopamine in regulating prefrontal function and working memory. In: Monsell S, Driver J, editors. Attention and performance XVIII; Control of cognitive processes. Cambridge, MA: MIT; 2000. pp. 713–737.
-
- Brett M, Anton JL, Valabregue R, Poline JB. Region of interest analysis using an SPM toolbox. Presented at the 8th International Conference on Functional Mapping of the Human Brain; June 2–6; Sendai, Japan. 2002.
-
- Bundesen C. A theory of visual-attention. Psychol Rev. 1990;97:523–547. - PubMed
-
- Bunzeck N, Düzel E. Absolute coding of stimulus novelty in the human substantia nigra/VTA. Neuron. 2006;51:369–379. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources