

Molecular architecture of the Goodpasture autoantigen in anti-GBM nephritis
- PMID: 20660402
- PMCID: PMC4144421
- DOI: 10.1056/NEJMoa0910500
Molecular architecture of the Goodpasture autoantigen in anti-GBM nephritis
Abstract
Background: In Goodpasture's disease, circulating autoantibodies bind to the noncollagenous-1 (NC1) domain of type IV collagen in the glomerular basement membrane (GBM). The specificity and molecular architecture of epitopes of tissue-bound autoantibodies are unknown. Alport's post-transplantation nephritis, which is mediated by alloantibodies against the GBM, occurs after kidney transplantation in some patients with Alport's syndrome. We compared the conformations of the antibody epitopes in Goodpasture's disease and Alport's post-transplantation nephritis with the intention of finding clues to the pathogenesis of anti-GBM glomerulonephritis.
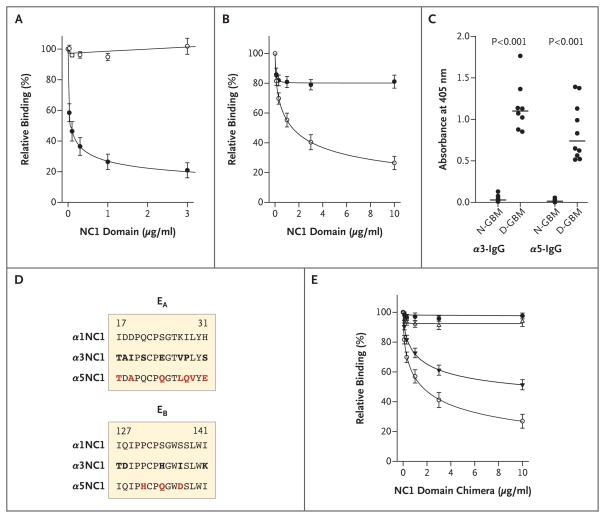
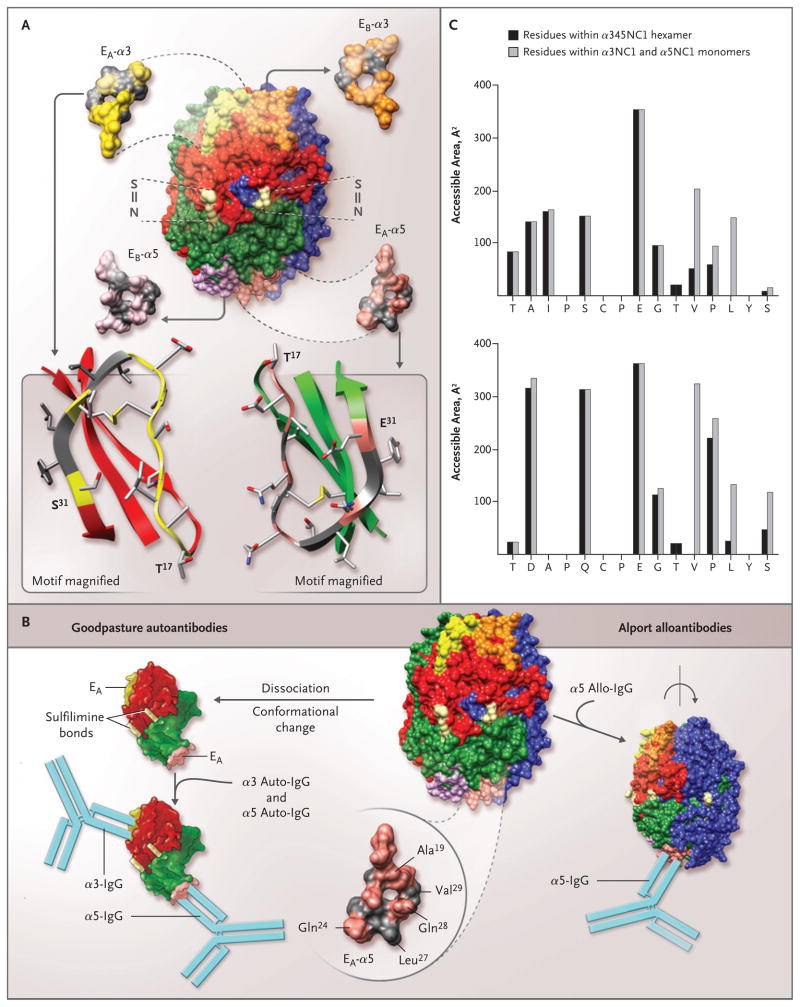
Methods: We used an enzyme-linked immunosorbent assay to determine the specificity of circulating autoantibodies and kidney-bound antibodies to NC1 domains. Circulating antibodies were analyzed in 57 patients with Goodpasture's disease, and kidney-bound antibodies were analyzed in 14 patients with Goodpasture's disease and 2 patients with Alport's post-transplantation nephritis. The molecular architecture of key epitope regions was deduced with the use of chimeric molecules and a three-dimensional model of the alpha345NC1 hexamer.
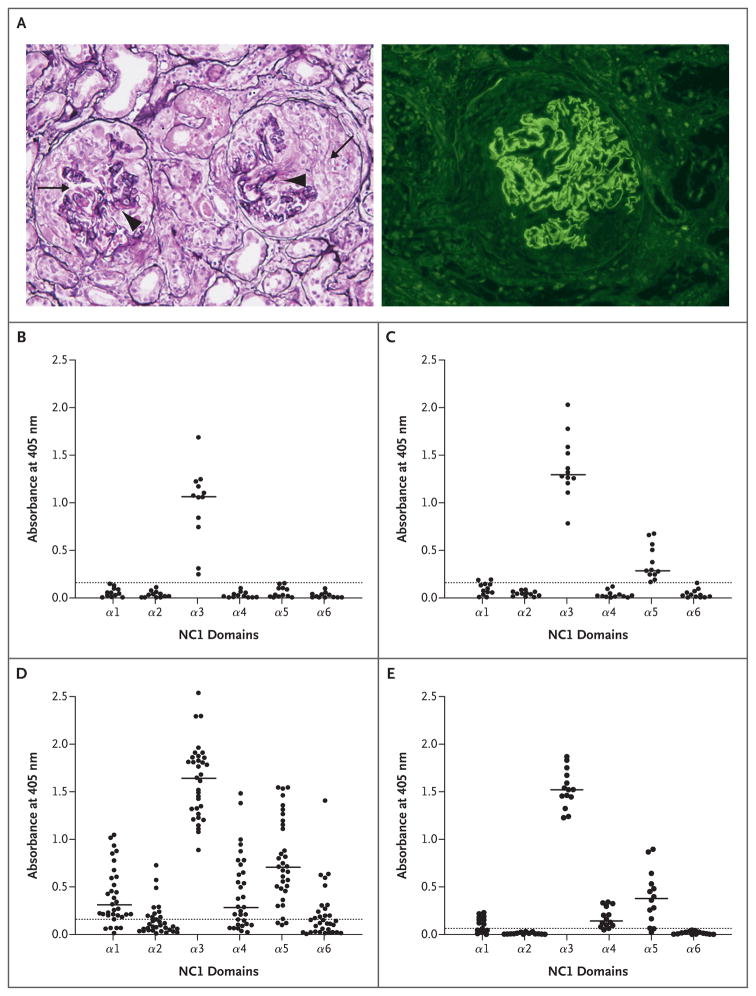
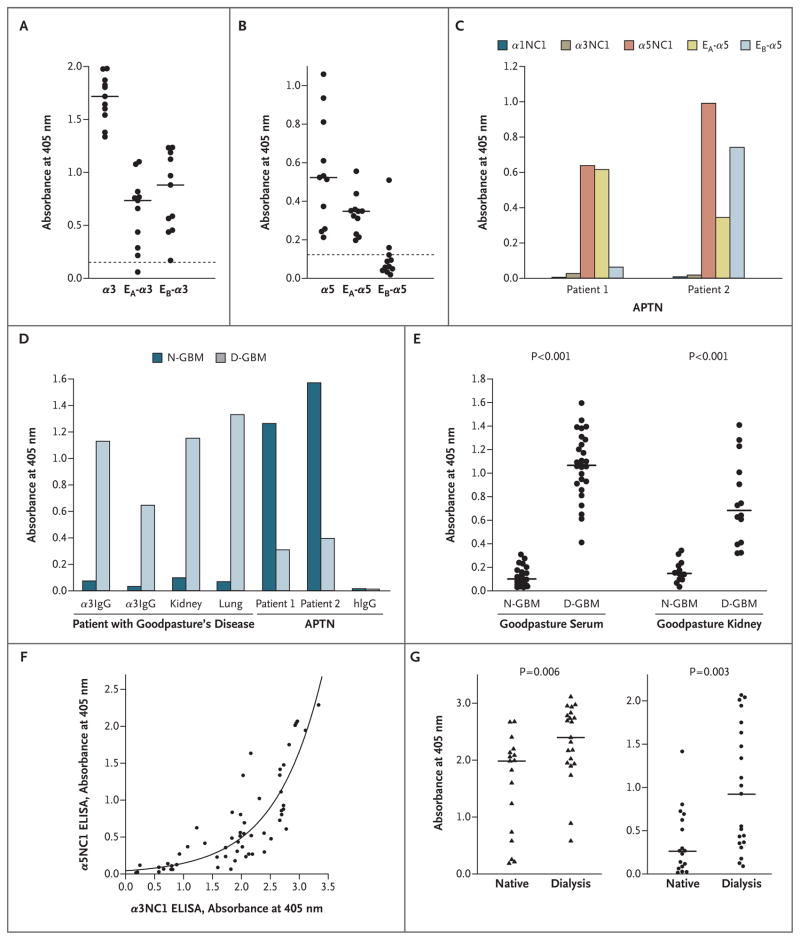
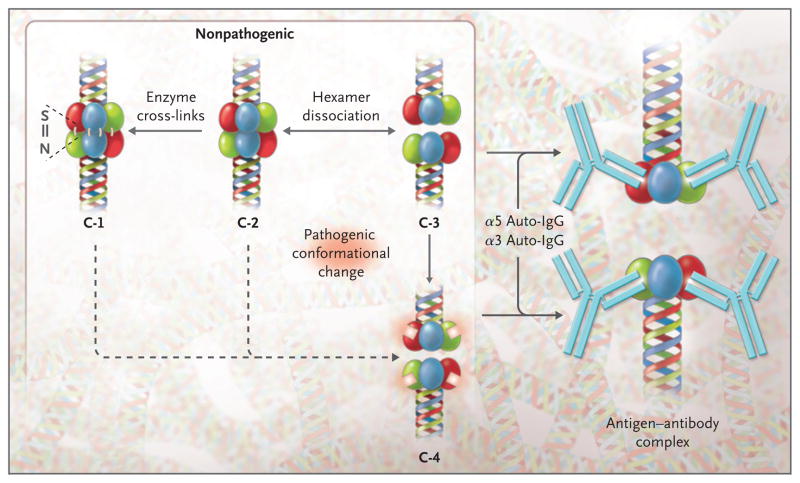
Results: In patients with Goodpasture's disease, both autoantibodies to the alpha3NC1 monomer and antibodies to the alpha5NC1 monomer (and fewer to the alpha4NC1 monomer) were bound in the kidneys and lungs, indicating roles for the alpha3NC1 and alpha5NC1 monomers as autoantigens. High antibody titers at diagnosis of anti-GBM disease were associated with ultimate loss of renal function. The antibodies bound to distinct epitopes encompassing region E(A) in the alpha5NC1 monomer and regions E(A) and E(B) in the alpha3NC1 monomer, but they did not bind to the native cross-linked alpha345NC1 hexamer. In contrast, in patients with Alport's post-transplantation nephritis, alloantibodies bound to the E(A) region of the alpha5NC1 subunit in the intact hexamer, and binding decreased on dissociation.
Conclusions: The development of Goodpasture's disease may be considered an autoimmune "conformeropathy" that involves perturbation of the quaternary structure of the alpha345NC1 hexamer, inducing a pathogenic conformational change in the alpha3NC1 and alpha5NC1 subunits, which in turn elicits an autoimmune response. (Funded by the National Institute of Diabetes and Digestive and Kidney Diseases.)
2010 Massachusetts Medical Society
Figures
Comment in
-
Goodpasture's disease--new secrets revealed.N Engl J Med. 2010 Jul 22;363(4):388-91. doi: 10.1056/NEJMe1006301. N Engl J Med. 2010. PMID: 20660408 No abstract available.
-
Molecular architecture of the Goodpasture autoantigen.N Engl J Med. 2010 Oct 28;363(18):1770; author reply 1771. doi: 10.1056/NEJMc1009688. N Engl J Med. 2010. PMID: 20979482 No abstract available.
References
-
- Wilson CB, Dixon FJ. Anti-glomerular basement membrane antibody-induced glomerulonephritis. Kidney Int. 1973;3:74–89. - PubMed
-
- Salama AD, Levy JB, Lightstone L, Pusey CD. Goodpasture’s disease. Lancet. 2001;358:917–20. - PubMed
-
- Saus J, Wieslander J, Langeveld JP, Quinones S, Hudson BG. Identification of the Goodpasture antigen as the alpha 3(IV) chain of collagen IV. J Biol Chem. 1988;263:13374–80. - PubMed
-
- Butkowski RJ, Langeveld JP, Wieslander J, Hamilton J, Hudson BG. Localization of the Goodpasture epitope to a novel chain of basement membrane collagen. J Biol Chem. 1987;262:7874–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources