

Ion channels set spike timing regularity of mammalian vestibular afferent neurons
- PMID: 20660422
- PMCID: PMC2957450
- DOI: 10.1152/jn.00396.2010
Ion channels set spike timing regularity of mammalian vestibular afferent neurons
Abstract
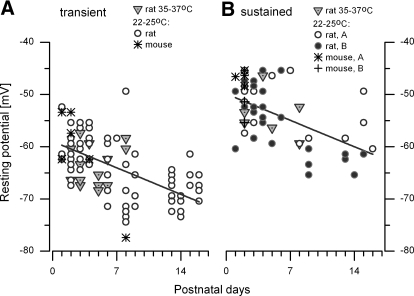
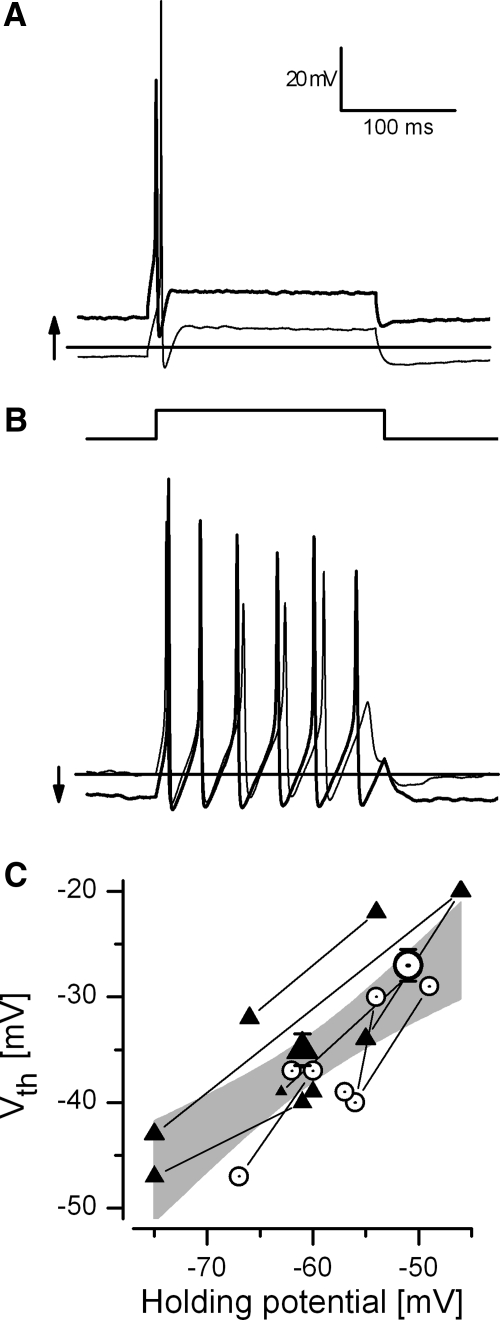
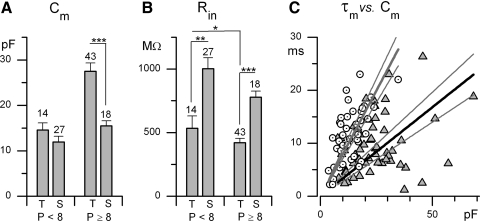
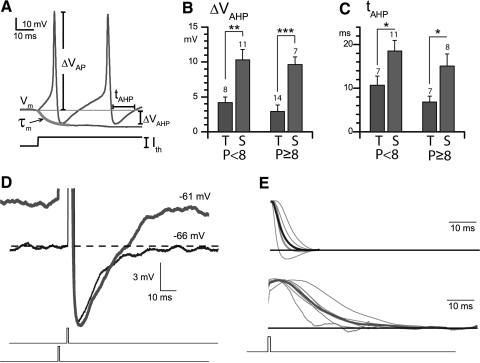
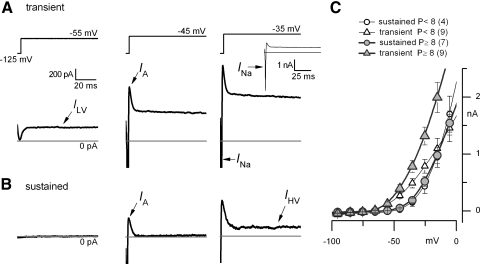
In the mammalian vestibular nerve, some afferents have highly irregular interspike intervals and others have highly regular intervals. To investigate whether spike timing is determined by the afferents' ion channels, we studied spiking activity in their cell bodies, isolated from the vestibular ganglia of young rats. Whole cell recordings were made with the perforated-patch method. As previously reported, depolarizing current steps revealed distinct firing patterns. Transient neurons fired one or two onset spikes, independent of current level. Sustained neurons were more heterogeneous, firing either trains of spikes or a spike followed by large voltage oscillations. We show that the firing pattern categories are robust, occurring at different temperatures and ages, both in mice and in rats. A difference in average resting potential did not cause the difference in firing patterns, but contributed to differences in afterhyperpolarizations. A low-voltage-activated potassium current (I(LV)) was previously implicated in the transient firing pattern. We show that I(LV) grew from the first to second postnatal week and by the second week comprised Kv1 and Kv7 (KCNQ) components. Blocking I(LV) converted step-evoked firing patterns from transient to sustained. Separated from their normal synaptic inputs, the neurons did not spike spontaneously. To test whether the firing-pattern categories might correspond to afferent populations of different regularity, we injected simulated excitatory postsynaptic currents at pseudorandom intervals. Sustained neurons responded to a given pattern of input with more regular firing than did transient neurons. Pharmacological block of I(LV) made firing more regular. Thus ion channel differences that produce transient and sustained firing patterns in response to depolarizing current steps can also produce irregular and regular spike timing.
Figures








References
-
- Adamson CL, Reid MA, Mo ZL, Bowne-English J, Davis RL. Firing features and potassium channel content of murine spiral ganglion neurons vary with cochlear location. J Comp Neurol 447: 331–350, 2002a - PubMed
-
- Baird RA, Desmadryl G, Fernández C, Goldberg JM. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J Neurophysiol 60: 182–203, 1988 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials