

doi: 10.1534/genetics.110.120717.
Epub 2010 Jul 26.
Targeting DNA double-strand breaks with TAL effector nucleases
Affiliations
- PMID: 20660643
- PMCID: PMC2942870
- DOI: 10.1534/genetics.110.120717
Item in Clipboard
Targeting DNA double-strand breaks with TAL effector nucleases
Genetics.
2010 Oct.
Abstract
Engineered nucleases that cleave specific DNA sequences in vivo are valuable reagents for targeted mutagenesis. Here we report a new class of sequence-specific nucleases created by fusing transcription activator-like effectors (TALEs) to the catalytic domain of the FokI endonuclease. Both native and custom TALE-nuclease fusions direct DNA double-strand breaks to specific, targeted sites.
Figures
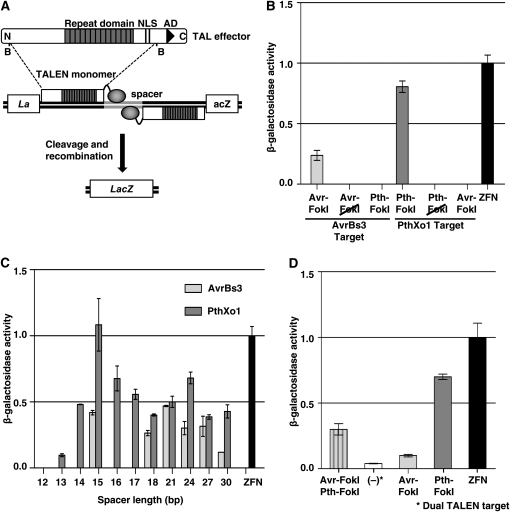
Structure and activity of TALE nucleases (TALENs). (A) Schematic of a transcription activator-like effector (TALE) protein. BamHI fragments were fused to the catalytic domain of the FokI endonuclease to create TALENs. TALEN activity was measured in an in vivo yeast assay that is described in the text (Townsend et al. 2009). N, N terminus; NLS, nuclear localization signal; B, BamHI site; AD, acidic activation domain. (B) Activity of TALENs constructed with TALEs AvrBs3 and PthXo1. Haploid cell types containing either TALEN expression or target plasmid in 200 μl of overnight culture were mated in YPD medium at 30°. After 4 hr, the YPD medium was replaced with 5 ml of selective medium and incubated overnight at 30°. Mated cultures were lysed, ONPG substrate added, and absorbance read at 415 nm using a 96-well plate reader (Townsend et al. 2009). β-Galactosidase levels were calculated as a function of substrate cleavage velocity and normalized to the ZFN positive control. Avr–FokI, AvrBs3 TALEN; Pth–FokI, PthXo1 TALEN; Avr–FokI and Pth–FokI, AvrBs3 and PthXo1 fusions to a catalytically inactive the version of FokI (Bitinaite et al. 1998); ZFN, zinc finger nuclease containing the Zif268 DNA binding domain as a positive control (Porteus and Baltimore 2003). (C) Activity of AvrBs3 and PthXo1 TALENS on targets with different spacer lengths. ZFN, Zif268-derived zinc finger nuclease. (D) Function of a heterodimeric TALEN. Activity in yeast containing PthXo1–FokI and AvrBs3–FokI expression vectors and a plasmid with a target consisting of recognition sites for each, in head-to-tail orientation separated by 15 bp is shown (Avr–FokI, Pth–FokI). Also shown for reference is activity of AvrBs3 (Avr–FokI) and PthXo1 (Pth–FokI) TALENS individually and Zif268 (ZFN) on their respective targets. As a negative control, a yeast culture with only the target site plasmid for Avr–FokI, Pth–FokI was assayed for LacZ activity [denoted as (−)].

Custom TALENs. To construct custom TALENs, repeats with the RVDs NI, HD, NN, and NG, specifying A, C, G, and T, respectively, were synthesized individually and assembled into modules of one, two, or three repeats. These modules were ligated sequentially into a derivative of the tal1c gene (Moscou and Bogdanove 2009) from which the original repeats had been removed. Five custom TALENs targeted to ADH1 from Arabidopsis and the zebrafish gridlock gene were created. (A) The RVD sequences of individual custom TALENs and their respective DNA recognition sequences are shown. (B) The yeast in vivo assay was used to determine the activity of custom TALENs. Plasmids containing two recognition sites for the respective TALEN in head-to-tail orientation separated by 16- to 18-bp spacers were used as targets. We chose spacer lengths on the basis of the distance closest to 15 bp from the 3′ end of the next neighboring (and opposing) candidate site, since we anticipated constructing heterodimeric TALENS that recognize native chromosomal sequences. Sixteen-base pair spacers were used for ADH1-360-12, ADH1-408-12r and 18-bp spacers for ADH1-928-12, ADH1-975-12r, and gridlock-2356-13r. The yeast assay was performed as described in Fig. 1B. (−), negative control with target site plasmids only; ZFN, zinc finger nuclease positive control.
References
-
- Boch, J., H. Scholze, S. Schornack, A. Landgraf, S. Hahn et al., 2009. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326 1509–1512. - PubMed
-
- Bogdanove, A. J., S. Schornack and T. Lahaye, 2010. TAL effectors: finding plant genes for disease and defense. Curr. Opin. Plant Biol. 13 394–401. - PubMed
-
- Bonas, U., R. E. Stall and B. Staskawicz, 1989. Genetic and structural characterization of the avirulence gene avrBs3 from Xanthomonas campestris pv. vesicatoria. Mol. Gen. Genet. 218 127–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
