

Review
doi: 10.1534/genetics.110.119297.
Dictyostelium finds new roles to model
Affiliations
- PMID: 20660652
- PMCID: PMC2907197
- DOI: 10.1534/genetics.110.119297
Item in Clipboard
Review
Dictyostelium finds new roles to model
Genetics.
2010 Jul.
Abstract
Any established or aspiring model organism must justify itself using two criteria: does the model organism offer experimental advantages not offered by competing systems? And will any discoveries made using the model be of wider relevance? This review addresses these issues for the social amoeba Dictyostelium and highlights some of the organisms more recent applications. These cover a remarkably wide gamut, ranging from sociobiological to medical research with much else in between.
Figures
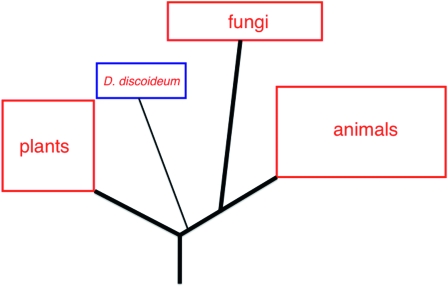
The evolutionary relationships of D. discoideum. This schematic, deduced from sequence comparisons, displays the evolutionary position of D. discoideum relative to other selected phyla.
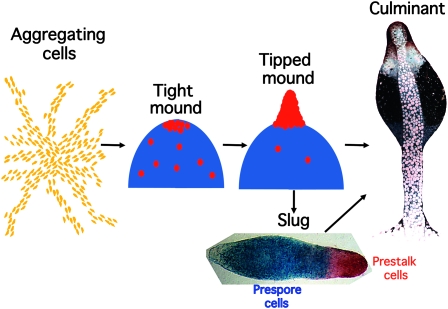
The Dictyostelium life cycle. Development in Dictyostelium is highly regulative and, depending on the local cell density, from 100 to 100,000 cells may aggregate together in response to cAMP signals emanating from the center of an aggregation territory. At the end of aggregation a mound of cells is formed and prespore cell differentiation, represented by blue shading, is induced by cAMP signaling. A subset of the uncommitted cells become prestalk cells (red circles) and migrate to the apex of the mound. There these cells form themselves into a nipple-shaped structure that extends to form an upright slug-shaped structure. Depending on environmental conditions, this structure may enter culmination to form a fruiting body immediately or may migrate for a time before completing development. The slug displayed was transformed with an ecmA-to-lacZ promoter fusion that is expressed in all prestalk cells (red) and a pspA-to-gus promoter fusion that is expressed in all prespore cells (blue). The two expression patterns were revealed by double enzymatic, gus–gal staining. The fruiting body is an electron microscopy image (copyright by M. J. Grimson and R. L. Blanton) that is false colored to show the position of the prestalk cells (red), upper cup cells (white), and spore cell precursors (blue).
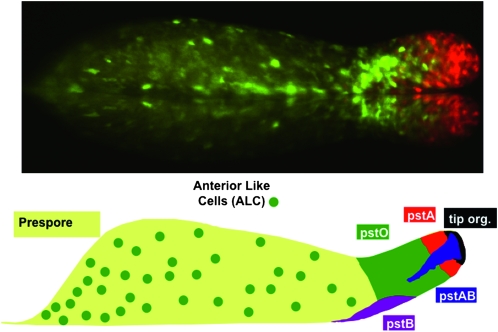
Prestalk and stalk cell heterogeneity. (Top) The slug displayed was transformed with two different ecmA promoter-to-GFP fusions that are expressed in pstA cells (red), pstO cells, or anterior-like cells (ALCs) (green) (D. Dormann , N. Zhukovskaya , J. G. Williams and C. J. Weijer , unpublished results). (Bottom) A representation of the different prestalk cell populations present in the slug. The anterior prestalk region contains four partially overlapping populations (Gaudet et al. 2008; http://dictybase.org/Dicty_Info/dicty_anatomy_ontology.html ). The pstA and pstO cells, respectively, occupy the front and rear halves and are identified by their ability to use different parts of the promoter of the ecmA gene. The pstAB cells are pstA cells that prematurely and sporadically express the ecmB gene. This is the same transitional event that occurs continuously during culmination. The pstAB cells sporadically drop backward and fall out the back of the migrating slug. At culmination, they form the inner part of the basal disc. The tip organizer cells are characterized by their ability to use a specific region of the promoter of the cudA gene. They control the migration and maintain the integrity of the slug. The pstB cells express ecmB at a higher level than ecmA and move backward and forward as a group along the anterior–posterior axis of the ventral surface of the prespore region. At culmination, they form the outer part of the basal disc. The ALCs are a mixed group of cells that are the direct precursors of the upper and lower cups of the culminant.

Degrees of anatomical complexity within the Dictyostelids. These are images of three of the species for which a complete genome sequence is available or soon will be available. Both A. subglobosum and P. pallidum are in group 2 of the molecular phylogeny of Schaap et al. (2006) while D. discoideum is in group 4. Note the differences in size, indicated by the scale bars. These images were kindly provided by F. Spiegel.
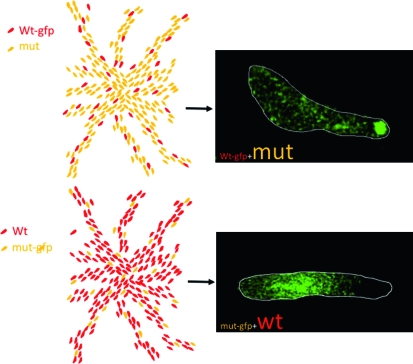
A typical synergy experiment as used in cheater assays. The drawings at the left represent aggregation territories formed by (top) a minority (usually 5–10%) of GFP-labeled parental cells mixed with a large excess of GFP-labeled cheater mutant cells and (bottom) a majority of unlabeled parental cells mixed with a minority of GFP-labeled cheater mutant cells. The images at the right are from such an experiment using a mutant defective in several aspects of late development (C. Sugden and J. Williams , unpublished data). In both variants of the mixing protocol, the mutant cells are selectively excluded from the prestalk zone.
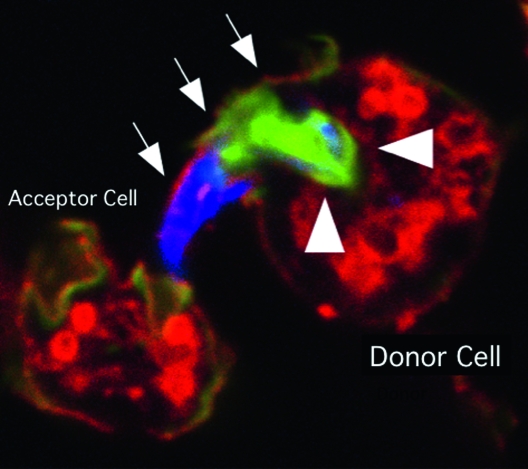
An ejectosome produced by a Dictyostelium cell infected with Mycobacterium marinum. A bundle of mycobacteria is being ejected from a donor cell via an ejectosome (Hagedorn et al. 2009). The plasma membrane bulge (arrows) is ruptured at the tip, where it contacts the acceptor cell. Actin tails, stained green with phalloidin (arrowheads), are polarized at the posterior of the bacteria, which are stained blue. This image was kindly provided by T. Soldati.
References
-
- Beug, H., G. Gerisch, S. Kempff, V. Riedel and G. Cremer, 1970. Specific inhibition of cell contact formation in Dictyostelium by univalent antibodies. Exp. Cell Res. 63 147–158. - PubMed
-
- Blaauw, M., M. H. K. Linskens and P. J. M. van Haastert, 2000. Efficient control of gene expression by a tetracycline-dependent transactivator in single Dictyostelium discoideum cells. Gene 252 71–82. - PubMed
-
- Bozzaro, S., C. Bucci and M. Steinert, 2008. Phagocytosis and host-pathogen interactions in Dictyostelium with a look at macrophages. Int. Rev. Cell Mol. Biol. 271 253–300. - PubMed
-
- Bromberg, J., and J. E. Darnell, Jr., 2000. The role of STATs in transcriptional control and their impact on cellular function. Oncogene 19 2468–2473. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
