

Modification of hERG1 channel gating by Cd2+
- PMID: 20660661
- PMCID: PMC2912066
- DOI: 10.1085/jgp.201010450
Modification of hERG1 channel gating by Cd2+
Abstract
Each of the four subunits in a voltage-gated potassium channel has a voltage sensor domain (VSD) that is formed by four transmembrane helical segments (S1-S4). In response to changes in membrane potential, intramembrane displacement of basic residues in S4 produces a gating current. As S4 moves through the membrane, its basic residues also form sequential electrostatic interactions with acidic residues in immobile regions of the S2 and S3 segments. Transition metal cations interact with these same acidic residues and modify channel gating. In human ether-á-go-go-related gene type 1 (hERG1) channels, Cd(2+) coordinated by D456 and D460 in S2 and D509 in S3 induces a positive shift in the voltage dependence of activation of ionic currents. Here, we characterize the effects of Cd(2+) on hERG1 gating currents in Xenopus oocytes using the cut-open Vaseline gap technique. Cd(2+) shifted the half-point (V(1/2)) for the voltage dependence of the OFF gating charge-voltage (Q(OFF)-V) relationship with an EC(50) of 171 microM; at 0.3 mM, V(1/2) was shifted by +50 mV. Cd(2+) also induced an as of yet unrecognized small outward current (I(Cd-out)) upon repolarization in a concentration- and voltage-dependent manner. We propose that Cd(2+) and Arg residues in the S4 segment compete for interaction with acidic residues in S2 and S3 segments, and that the initial inward movement of S4 associated with membrane repolarization displaces Cd(2+) in an outward direction to produce I(Cd-out). Co(2+), Zn(2+), and La(3+) at concentrations that caused approximately +35-mV shifts in the Q(OFF)-V relationship did not induce a current similar to I(Cd-out), suggesting that the binding site for these cations or their competition with basic residues in S4 differs from Cd(2+). New Markov models of hERG1 channels were developed that describe gating currents as a noncooperative two-phase process of the VSD and can account for changes in these currents caused by extracellular Cd(2+).
Figures
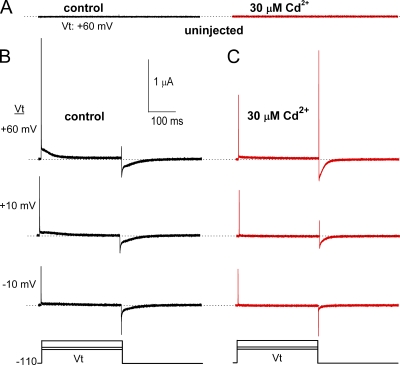




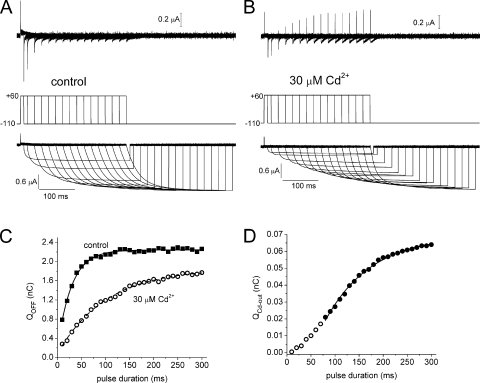








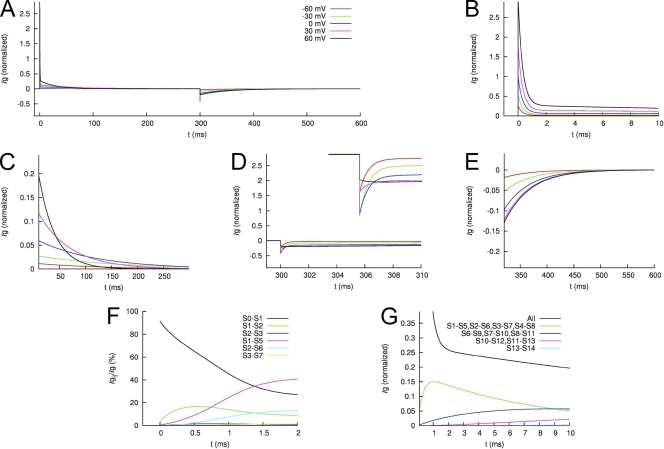



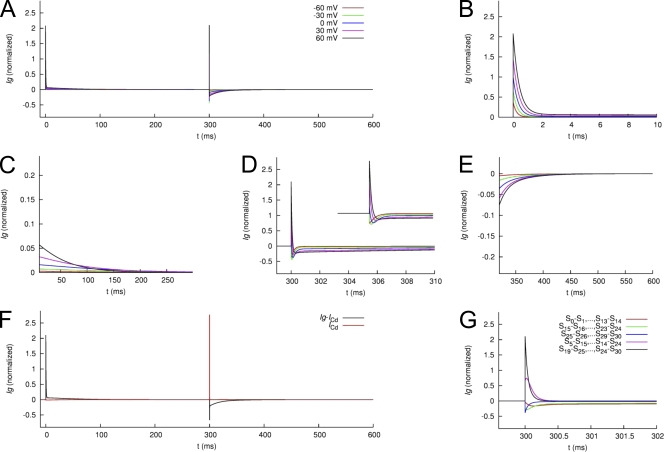

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
