

B cell activating factor (BAFF) and T cells cooperate to breach B cell tolerance in lupus-prone New Zealand Black (NZB) mice
- PMID: 20661465
- PMCID: PMC2908288
- DOI: 10.1371/journal.pone.0011691
B cell activating factor (BAFF) and T cells cooperate to breach B cell tolerance in lupus-prone New Zealand Black (NZB) mice
Abstract
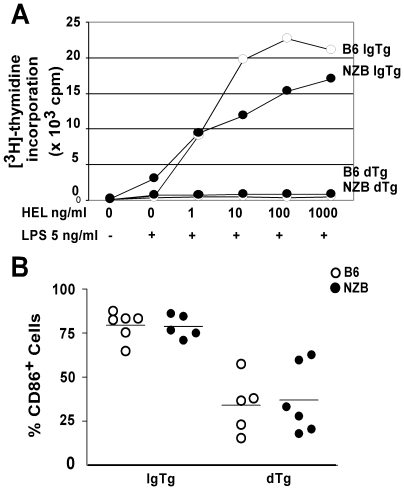
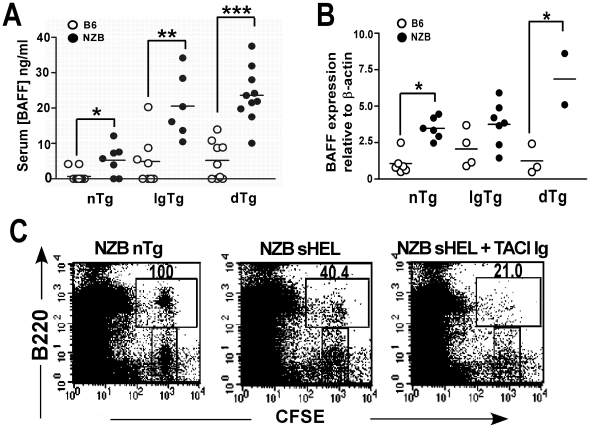
The presence of autoantibodies in New Zealand Black (NZB) mice suggests a B cell tolerance defect however the nature of this defect is unknown. To determine whether defects in B cell anergy contribute to the autoimmune phenotype in NZB mice, soluble hen egg lysozyme (sHEL) and anti-HEL Ig transgenes were bred onto the NZB background to generate double transgenic (dTg) mice. NZB dTg mice had elevated levels of anti-HEL antibodies, despite apparently normal B cell functional anergy in-vitro. NZB dTg B cells also demonstrated increased survival and abnormal entry into the follicular compartment following transfer into sHEL mice. Since this process is dependent on BAFF, BAFF serum and mRNA levels were assessed and were found to be significantly elevated in NZB dTg mice. Treatment of NZB sHEL recipient mice with TACI-Ig reduced NZB dTg B cell survival following adoptive transfer, confirming the role of BAFF in this process. Although NZB mice had modestly elevated BAFF, the enhanced NZB B cell survival response appeared to result from an altered response to BAFF. In contrast, T cell blockade had a minimal effect on B cell survival, but inhibited anti-HEL antibody production. The findings suggest that the modest BAFF elevations in NZB mice are sufficient to perturb B cell tolerance, particularly when acting in concert with B cell functional abnormalities and T cell help.
Conflict of interest statement
Figures



Similar articles
-
Multiple tolerance defects contribute to the breach of B cell tolerance in New Zealand Black chromosome 1 congenic mice.PLoS One. 2017 Jun 19;12(6):e0179506. doi: 10.1371/journal.pone.0179506. eCollection 2017. PLoS One. 2017. PMID: 28628673 Free PMC article.
-
Autoreactive B cells in lupus-prone New Zealand black mice exhibit aberrant survival and proliferation in the presence of self-antigen in vivo.J Immunol. 2004 Feb 1;172(3):1553-60. doi: 10.4049/jimmunol.172.3.1553. J Immunol. 2004. PMID: 14734734
-
Altered toll-like receptor responsiveness underlies a dominant heritable defect in B cell tolerance in autoimmune New Zealand Black mice.Eur J Immunol. 2018 Mar;48(3):492-497. doi: 10.1002/eji.201747287. Epub 2018 Jan 19. Eur J Immunol. 2018. PMID: 29251774 Free PMC article.
-
B cells flying solo.Immunol Cell Biol. 2008 Jan;86(1):40-6. doi: 10.1038/sj.icb.7100142. Immunol Cell Biol. 2008. PMID: 18172443 Review.
-
Does the BAFF dysregulation play a major role in the pathogenesis of systemic lupus erythematosus?J Autoimmun. 2008 Feb-Mar;30(1-2):63-7. doi: 10.1016/j.jaut.2007.11.001. Epub 2007 Dec 21. J Autoimmun. 2008. PMID: 18155417 Review.
Cited by
-
Regulation of basement membrane-reactive B cells in BXSB, (NZBxNZW)F1, NZB, and MRL/lpr lupus mice.Autoimmunity. 2013 May;46(3):188-204. doi: 10.3109/08916934.2012.746671. Epub 2013 Jan 9. Autoimmunity. 2013. PMID: 23157336 Free PMC article.
-
Intrinsic autoimmune capacities of hematopoietic cells from female New Zealand hybrid mice.Genes Immun. 2014 Apr-May;15(3):153-61. doi: 10.1038/gene.2014.2. Epub 2014 Jan 30. Genes Immun. 2014. PMID: 24477163 Free PMC article.
-
Requirement for Transcription Factor Ets1 in B Cell Tolerance to Self-Antigens.J Immunol. 2015 Oct 15;195(8):3574-83. doi: 10.4049/jimmunol.1500776. Epub 2015 Sep 9. J Immunol. 2015. PMID: 26355157 Free PMC article.
-
Multiple tolerance defects contribute to the breach of B cell tolerance in New Zealand Black chromosome 1 congenic mice.PLoS One. 2017 Jun 19;12(6):e0179506. doi: 10.1371/journal.pone.0179506. eCollection 2017. PLoS One. 2017. PMID: 28628673 Free PMC article.
-
Lack of T cells in Act1-deficient mice results in elevated IgM-specific autoantibodies but reduced lupus-like disease.Eur J Immunol. 2012 Jul;42(7):1695-705. doi: 10.1002/eji.201142238. Eur J Immunol. 2012. PMID: 22585710 Free PMC article.
References
-
- Theofilopoulos AN, Dixon FJ. Murine models of systemic lupus erythematosus. Adv Immunol. 1985;37:269–390. - PubMed
-
- Izui S, McConahey PJ, Dixon FJ. Increased spontaneous polyclonal activation of B lymphocytes in mice with spontaneous autoimmune disease. J Immunol. 1978;121:2213–9. - PubMed
-
- Wither JE, Roy V, Brennan LA. Activated B cells express increased levels of costimulatory molecules in young autoimmune NZB and (NZB x NZW)F(1) mice. Clin Immunol. 2000;94:51–63. - PubMed
-
- Manny N, Datta SK, Schwartz RS. Synthesis of IgM by cells of NZB and SWR mice and their crosses. J Immunol. 1979;122:1220–7. - PubMed
-
- Moutsopoulos HM, Boehm-Truitt M, Kassan SS, Chused TM. Demonstration of activation of B lymphocytes in New Zealand black mice at birth by an immunoradiometric assay for murine IgM. J Immunol. 1977;119:1639–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
