

Uropathogenic Escherichia coli modulates immune responses and its curli fimbriae interact with the antimicrobial peptide LL-37
- PMID: 20661475
- PMCID: PMC2908543
- DOI: 10.1371/journal.ppat.1001010
Uropathogenic Escherichia coli modulates immune responses and its curli fimbriae interact with the antimicrobial peptide LL-37
Abstract
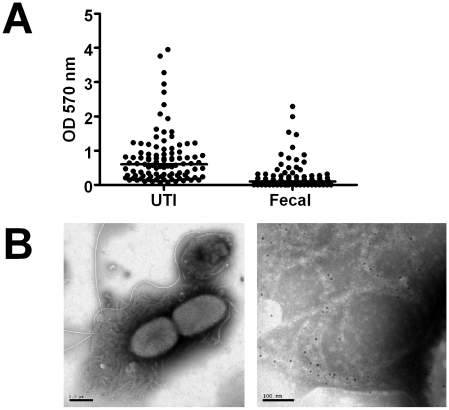
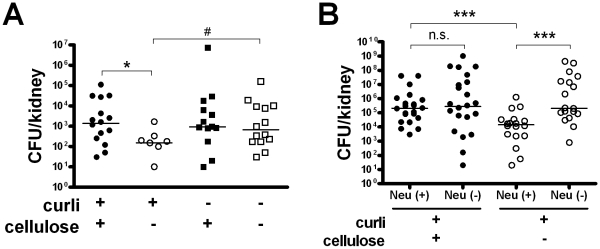
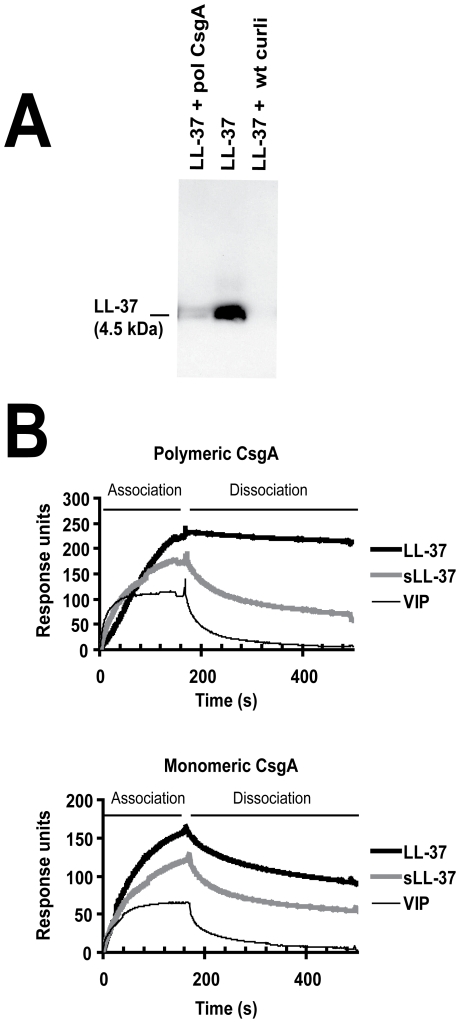
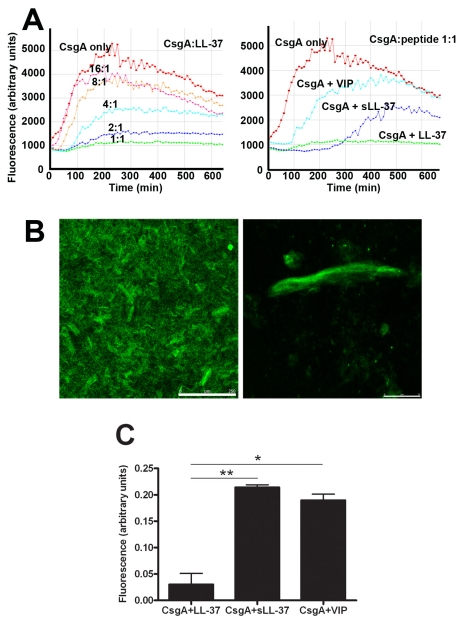
Bacterial growth in multicellular communities, or biofilms, offers many potential advantages over single-cell growth, including resistance to antimicrobial factors. Here we describe the interaction between the biofilm-promoting components curli fimbriae and cellulose of uropathogenic E. coli and the endogenous antimicrobial defense in the urinary tract. We also demonstrate the impact of this interplay on the pathogenesis of urinary tract infections. Our results suggest that curli and cellulose exhibit differential and complementary functions. Both of these biofilm components were expressed by a high proportion of clinical E. coli isolates. Curli promoted adherence to epithelial cells and resistance against the human antimicrobial peptide LL-37, but also increased the induction of the proinflammatory cytokine IL-8. Cellulose production, on the other hand, reduced immune induction and hence delayed bacterial elimination from the kidneys. Interestingly, LL-37 inhibited curli formation by preventing the polymerization of the major curli subunit, CsgA. Thus, even relatively low concentrations of LL-37 inhibited curli-mediated biofilm formation in vitro. Taken together, our data demonstrate that biofilm components are involved in the pathogenesis of urinary tract infections by E. coli and can be a target of local immune defense mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Discovery of New Genes Involved in Curli Production by a Uropathogenic Escherichia coli Strain from the Highly Virulent O45:K1:H7 Lineage.mBio. 2018 Aug 21;9(4):e01462-18. doi: 10.1128/mBio.01462-18. mBio. 2018. PMID: 30131362 Free PMC article.
-
Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli.Environ Microbiol. 2009 Apr;11(4):992-1006. doi: 10.1111/j.1462-2920.2008.01824.x. Epub 2009 Nov 14. Environ Microbiol. 2009. PMID: 19187284 Free PMC article.
-
Impact of biofilm matrix components on interaction of commensal Escherichia coli with the gastrointestinal cell line HT-29.Cell Mol Life Sci. 2006 Oct;63(19-20):2352-63. doi: 10.1007/s00018-006-6222-4. Cell Mol Life Sci. 2006. PMID: 16952050 Free PMC article.
-
A systematic review and meta-analysis of antibiotic resistance patterns, and the correlation between biofilm formation with virulence factors in uropathogenic E. coli isolated from urinary tract infections.Microb Pathog. 2020 Jul;144:104196. doi: 10.1016/j.micpath.2020.104196. Epub 2020 Apr 10. Microb Pathog. 2020. PMID: 32283258
-
Virulence factors of uropathogenic E. coli and their interaction with the host.Adv Microb Physiol. 2014;65:337-72. doi: 10.1016/bs.ampbs.2014.08.006. Epub 2014 Nov 4. Adv Microb Physiol. 2014. PMID: 25476769 Review.
Cited by
-
Bacterial resistance to antibacterial agents: Mechanisms, control strategies, and implications for global health.Sci Total Environ. 2023 Feb 20;860:160461. doi: 10.1016/j.scitotenv.2022.160461. Epub 2022 Nov 23. Sci Total Environ. 2023. PMID: 36435256 Free PMC article. Review.
-
An optotracer-based antibiotic susceptibility test specifically targeting the biofilm lifestyle of Salmonella.Biofilm. 2022 Sep 8;4:100083. doi: 10.1016/j.bioflm.2022.100083. eCollection 2022 Dec. Biofilm. 2022. PMID: 36117547 Free PMC article.
-
Amplifying renal immunity: the role of antimicrobial peptides in pyelonephritis.Nat Rev Nephrol. 2015 Nov;11(11):642-55. doi: 10.1038/nrneph.2015.105. Epub 2015 Jul 7. Nat Rev Nephrol. 2015. PMID: 26149835 Review.
-
The Potential of Human Peptide LL-37 as an Antimicrobial and Anti-Biofilm Agent.Antibiotics (Basel). 2021 May 29;10(6):650. doi: 10.3390/antibiotics10060650. Antibiotics (Basel). 2021. PMID: 34072318 Free PMC article. Review.
-
Whole-genome analysis of the colonization-resistant bacterium Phytobacter sp. SCO41T isolated from Bacillus nematocida B16-fed adult Caenorhabditis elegans.Mol Biol Rep. 2019 Apr;46(2):1563-1575. doi: 10.1007/s11033-018-04574-w. Epub 2019 Mar 16. Mol Biol Rep. 2019. PMID: 30879274
References
-
- Roberts RC, Mohr CD, Shapiro L. Developmental programs in bacteria. Curr Top Dev Biol. 1996;34:207–257. - PubMed
-
- Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Microbial biofilms. Annu Rev Microbiol. 1995;49:711–745. - PubMed
-
- Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Micro. 2004;2:95–108. - PubMed
-
- Shapiro JA. Thinking about bacterial populations as multicellular organisms. Ann Rev Microbiol. 1998;52:81–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
