

The molecular and gene regulatory signature of a neuron
- PMID: 20663572
- PMCID: PMC2947585
- DOI: 10.1016/j.tins.2010.05.006
The molecular and gene regulatory signature of a neuron
Abstract
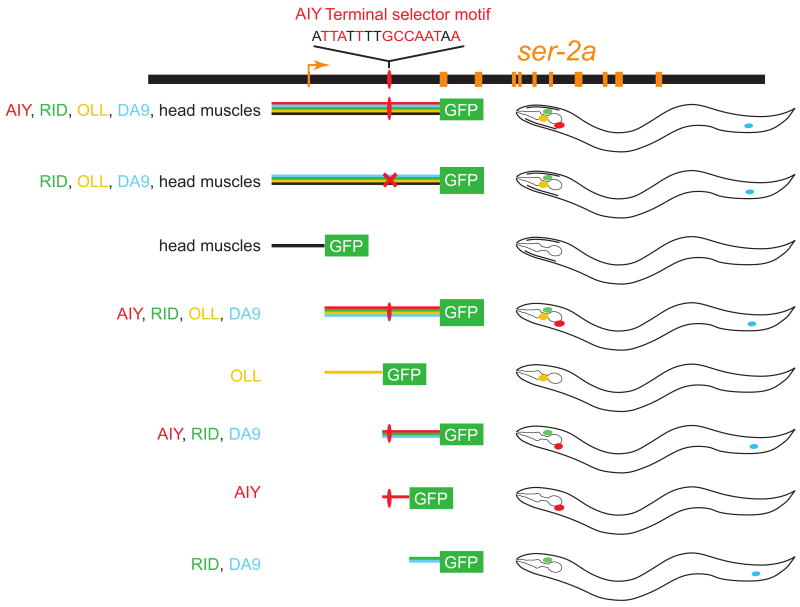
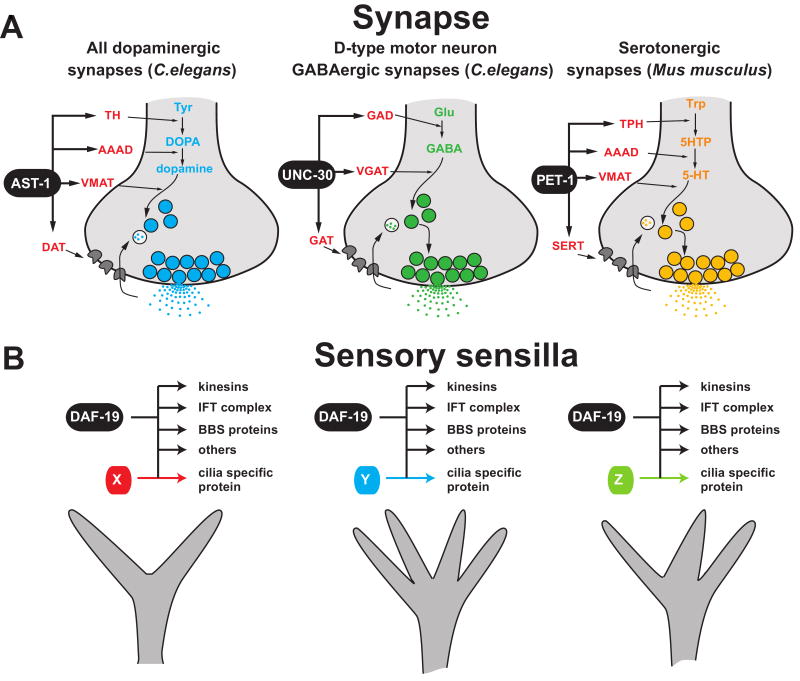
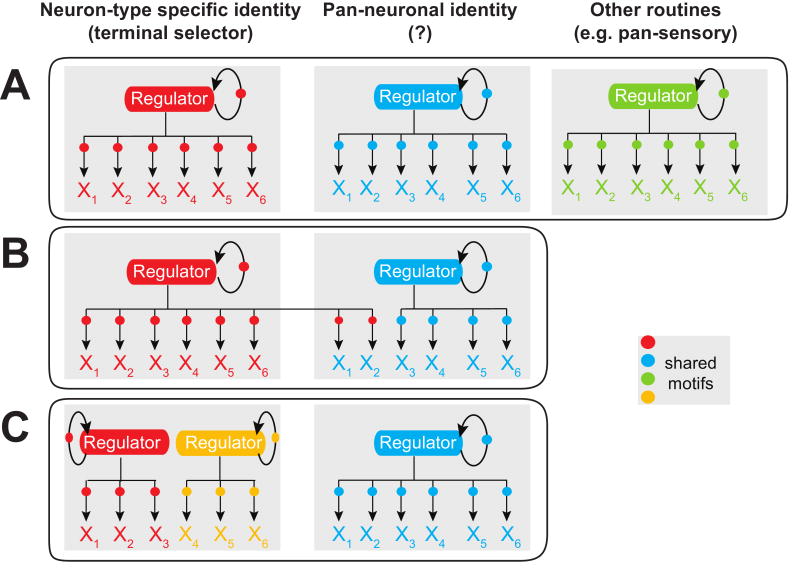
Neuron-type specific gene batteries define the morphological and functional diversity of cell types in the nervous system. Here, we discuss the composition of neuron-type specific gene batteries and illustrate gene regulatory strategies which determine the unique gene expression profiles and molecular composition of individual neuronal cell types from C. elegans to higher vertebrates. Based on principles learned from prokaryotic gene regulation, we argue that neuronal terminal gene batteries are functionally grouped into parallel-acting 'regulons'. The theoretical concepts discussed here provide testable hypotheses for future experimental analysis of the exact gene-regulatory mechanisms employed in the generation of neuronal diversity and identity.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures



References
-
- Lopez-Munoz F, Boya J, Alamo C. Neuron theory, the cornerstone of neuroscience, on the centenary of the Nobel Prize award to Santiago Ramon y Cajal. Brain Res Bull. 2006;70:391–405. - PubMed
-
- Waldeyer W. Ueber einige neuere Forschungen im Gebiete der Anatomie des Centralnervensystems. Deutsche Medicinische Wochenschrift. 1891;50:1352–1356.
-
- Garcia-Bellido A. Genetic control of wing disc development in Drosophila. Ciba Found Symp. 1975;29:161–182. - PubMed
-
- Mann RS, Carroll SB. Molecular mechanisms of selector gene function and evolution. Curr Opin Genet Dev. 2002;12:592–600. - PubMed
-
- Tansey EM. Not committing barbarisms: Sherrington and the synapse, 1897. Brain Res Bull. 1997;44:211–212. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
