

Cloning, functional characterization, and co-expression studies of a novel aquaporin (FaPIP2;1) of strawberry fruit
- PMID: 20663858
- PMCID: PMC2935871
- DOI: 10.1093/jxb/erq210
Cloning, functional characterization, and co-expression studies of a novel aquaporin (FaPIP2;1) of strawberry fruit
Abstract
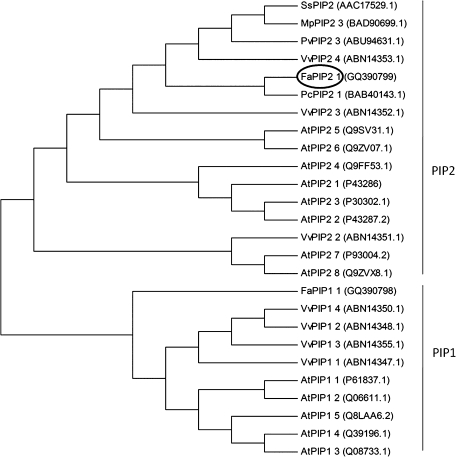
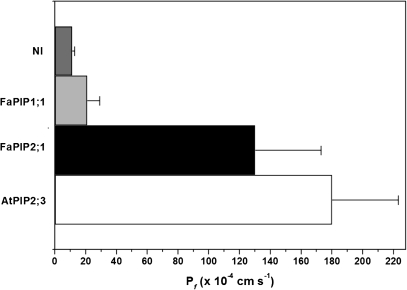
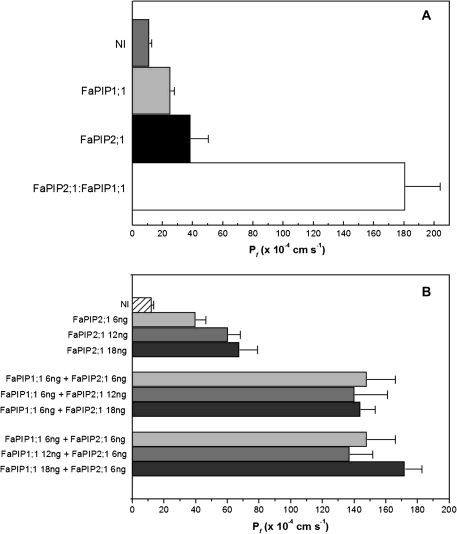
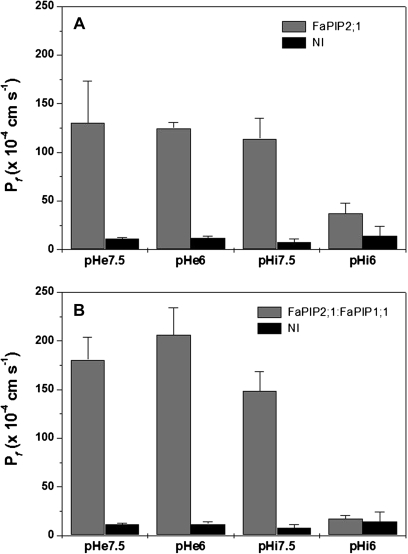
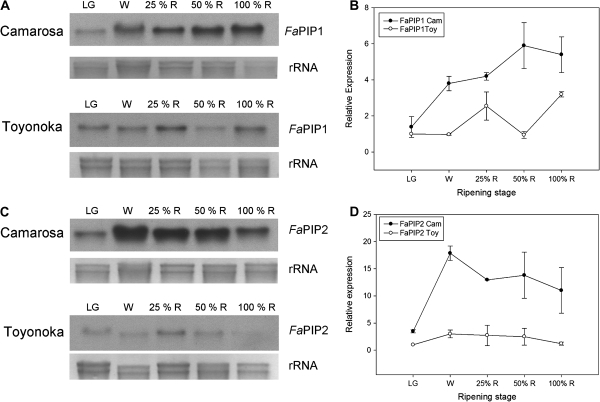
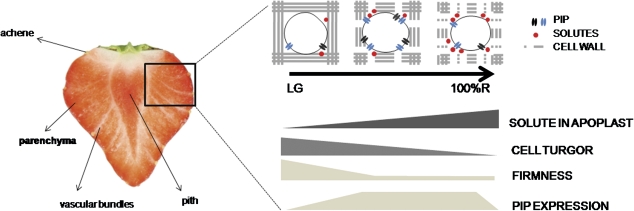
In strawberry, the putative participation of aquaporins should be considered during fruit ripening. Furthermore, the availability of different firmness cultivars in this non-climacteric fruit is a very useful tool to determine their involvement in softening. In a previous work, the cloning of a strawberry fruit-specific aquaporin, FaPIP1;1, which showed an expression profile associated with fruit ripening was reported. Here, FaPIP2;1, an aquaporin subtype of PIP2 was cloned and its functional characterization in Xenopus oocytes determined. The FaPIP2;1 gene encodes a water channel with high water permeability (P(f)) that is regulated by cytosolic pH. Interestingly, the co-expression of both FaPIP subtypes resulted in an enhancement of water permeability, showing P(f) values that exceeds their individual contribution. The expression pattern of both aquaporin subtypes in two cultivars with contrasting fruit firmness showed that the firmer cultivar (Camarosa) has a higher accumulation of FaPIP1 and FaPIP2 mRNAs during fruit ripening when compared with the softer cultivar (Toyonoka). In conclusion, not only FaPIP aquaporins showed an expression pattern associated with fruit firmness but it was also shown that the enhancement of water transfer through the plasma membrane is coupled to the presence/absence of the co-expression of both subtypes.
Figures

References
-
- Agre P, Mathai JC, Smith BL, Preston GM. Functional analyses of aquaporin water channel proteins. Methods in Enzymology. 1999;294:550–572. - PubMed
-
- Aharoni A, Keizer LCP, Van Den Broeck HC, Blanco-Portales R, Muñoz-Blanco J, Bois G, Smit P, De Vos RCH, O'Connell AP. Novel insight into vascular, stress, and auxin-dependent and -independent gene expression programs in strawberry, a nonclimacteric fruit. Plant Physiology. 2002;129:1019–1031. - PMC - PubMed
-
- Azad AK, Katsuhara M, Sawa Y, Ishikawa T, Shibata H. Characterization of four plasma membrane aquaporins in tulip petals: a putative homolog is regulated by phosphorylation. Plant and Cell Physiology. 2008;49:1196–1208. - PubMed
