

SUMOylation modulates the function of Aurora-B kinase
- PMID: 20663916
- PMCID: PMC2915883
- DOI: 10.1242/jcs.065565
SUMOylation modulates the function of Aurora-B kinase
Abstract
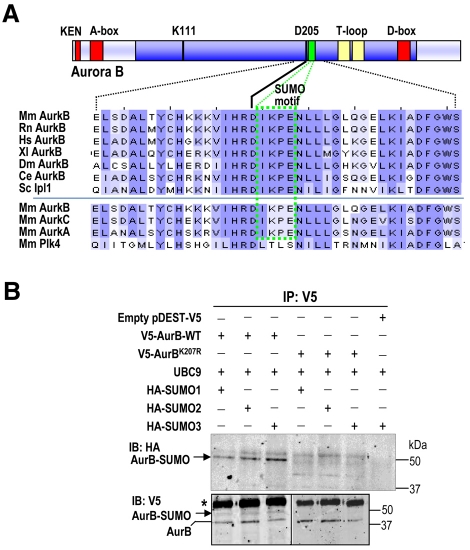
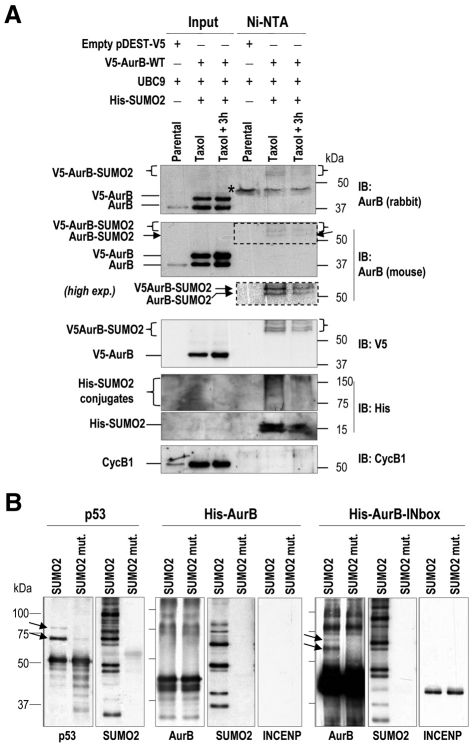
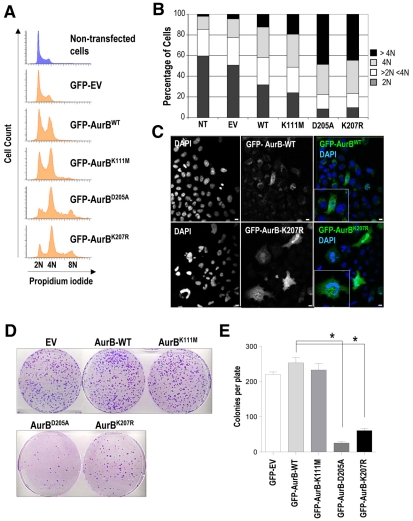
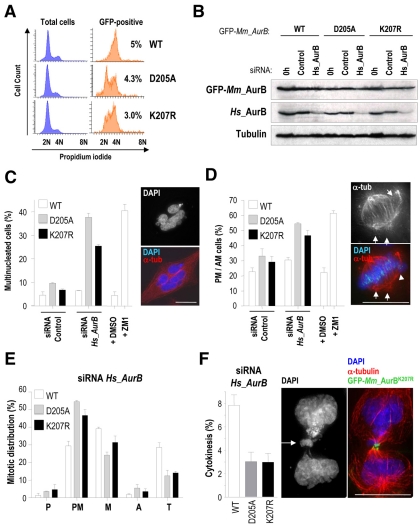
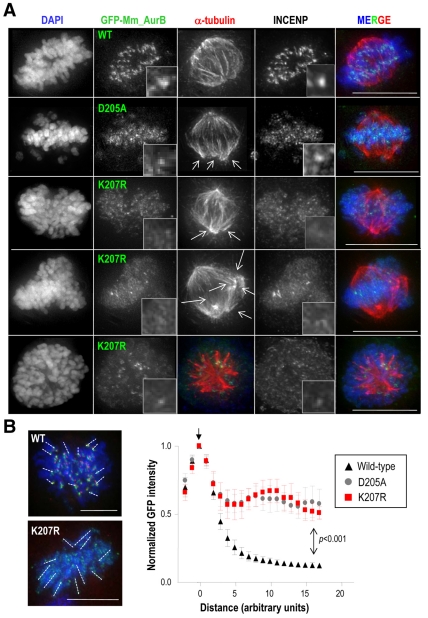
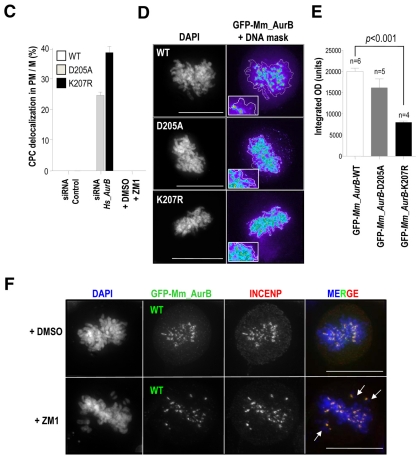
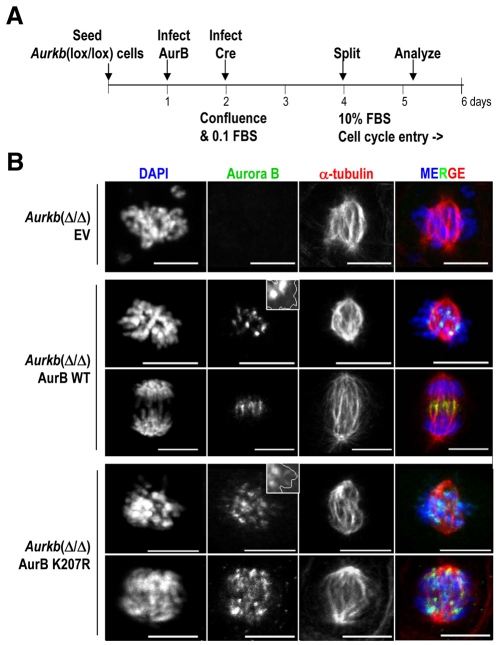
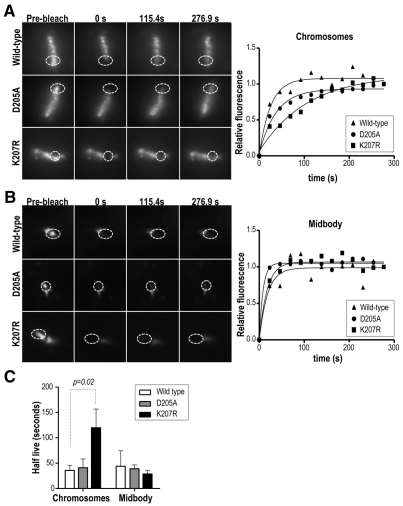
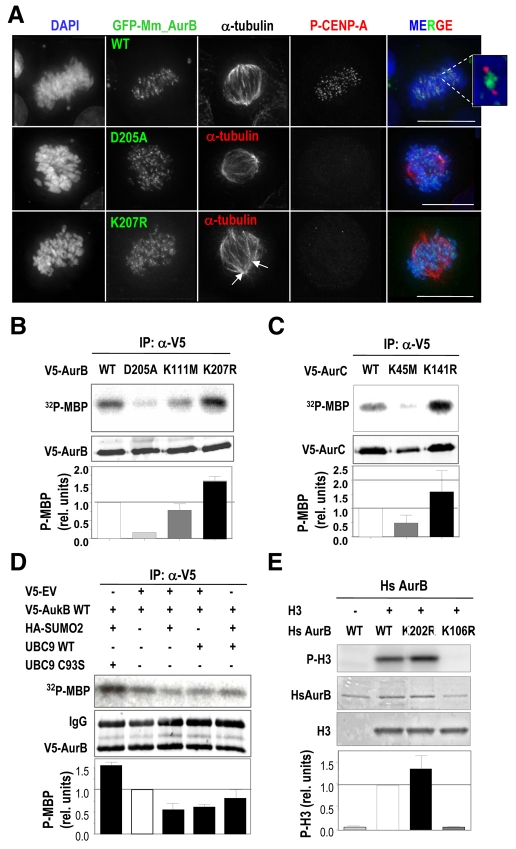
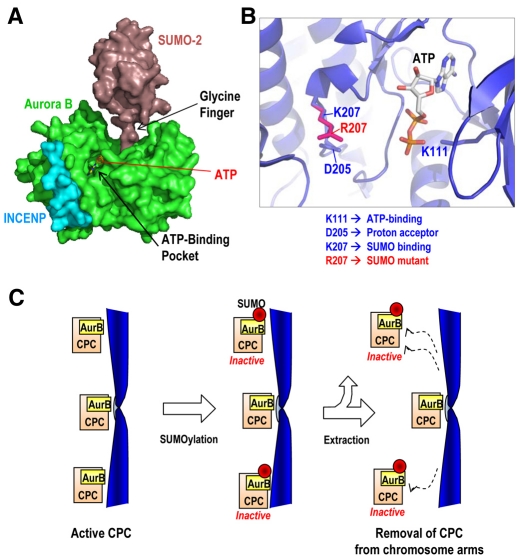
Aurora kinases are central regulators of mitotic-spindle assembly, chromosome segregation and cytokinesis. Aurora B is a member of the chromosomal passenger complex (CPC) with crucial functions in regulation of the attachment of kinetochores to microtubules and in cytokinesis. We report here that Aurora B contains a conserved SUMO modification motif within its kinase domain. Aurora B can bind SUMO peptides in vitro when bound to the IN-box domain of its CPC partner INCENP. Mutation of Lys207 to arginine (Aurora B(K207R)) impairs the formation of conjugates of Aurora B and SUMO in vivo. Expression of the SUMO-null form of Aurora B results in abnormal chromosome segregation and cytokinesis failure and it is not able to rescue mitotic defects in Aurora-B-knockout cells. These defects are accompanied by increased levels of the CPC on chromosome arms and defective centromeric function, as detected by decreased phosphorylation of the Aurora-B substrate CENP-A. The Aurora-B(K207R) mutant does not display reduced kinase activity, suggesting that functional defects are probably a consequence of the altered localization, rather than decreased intrinsic kinase activity. These data suggest that SUMOylation of Aurora B modulates its function, possibly by mediating the extraction of CPC complexes from chromosome arms during prometaphase.
Figures
References
-
- Adams R. R., Wheatley S. P., Gouldsworthy A. M., Kandels-Lewis S. E., Carmena M., Smythe C., Gerloff D. L., Earnshaw W. C. (2000). INCENP binds the Aurora-related kinase AIRK2 and is required to target it to chromosomes, the central spindle and cleavage furrow. Curr. Biol. 10, 1075-1078 - PubMed
-
- Azuma Y., Dasso M. (2002). A new clue at the nuclear pore: RanBP2 is an E3 enzyme for SUMO1. Dev. Cell 2, 130-131 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
