

Functional diversity of microbial decomposers facilitates plant coexistence in a plant-microbe-soil feedback model
- PMID: 20663953
- PMCID: PMC2922608
- DOI: 10.1073/pnas.0914281107
Functional diversity of microbial decomposers facilitates plant coexistence in a plant-microbe-soil feedback model
Abstract
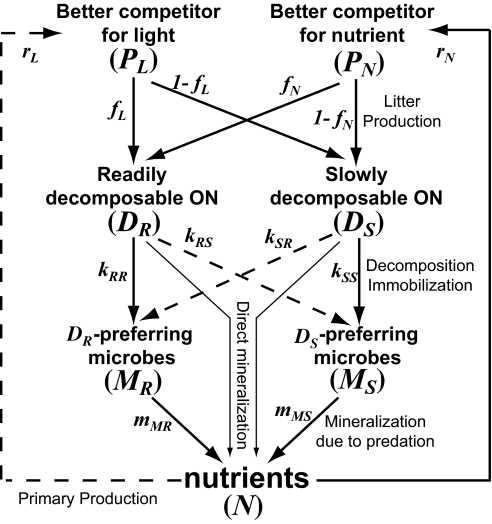
Theory and empirical evidence suggest that plant-soil feedback (PSF) determines the structure of a plant community and nutrient cycling in terrestrial ecosystems. The plant community alters the nutrient pool size in soil by affecting litter decomposition processes, which in turn shapes the plant community, forming a PSF system. However, the role of microbial decomposers in PSF function is often overlooked, and it remains unclear whether decomposers reinforce or weaken litter-mediated plant control over nutrient cycling. Here, we present a theoretical model incorporating the functional diversity of both plants and microbial decomposers. Two fundamental microbial processes are included that control nutrient mineralization from plant litter: (i) assimilation of mineralized nutrient into the microbial biomass (microbial immobilization), and (ii) release of the microbial nutrients into the inorganic nutrient pool (net mineralization). With this model, we show that microbial diversity may act as a buffer that weakens plant control over the soil nutrient pool, reversing the sign of PSF from positive to negative and facilitating plant coexistence. This is explained by the decoupling of litter decomposability and nutrient pool size arising from a flexible change in the microbial community composition and decomposition processes in response to variations in plant litter decomposability. Our results suggest that the microbial community plays a central role in PSF function and the plant community structure. Furthermore, the results strongly imply that the plant-centered view of nutrient cycling should be changed to a plant-microbe-soil feedback system, by incorporating the community ecology of microbial decomposers and their functional diversity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
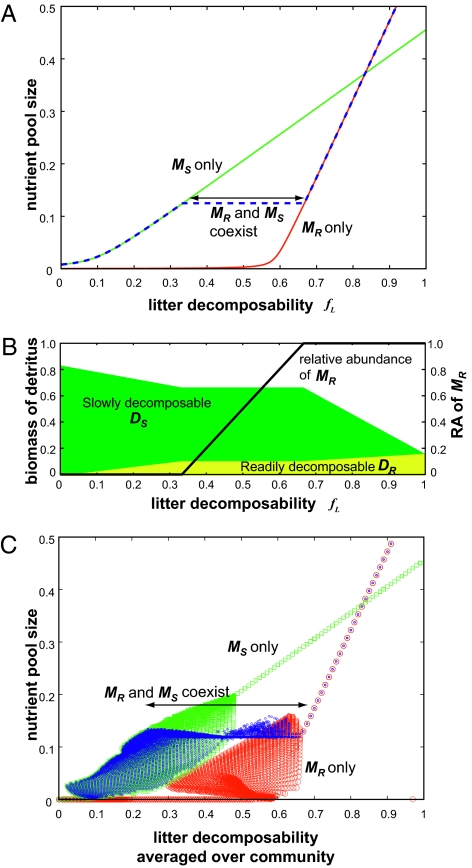
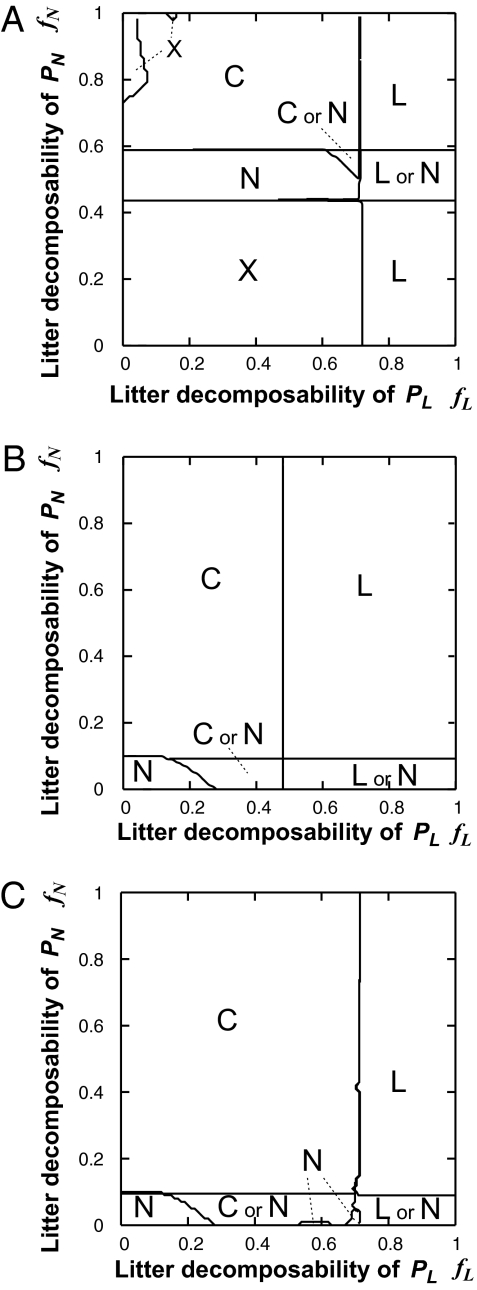
 , from t = 45,000 to t = 50,000) and the average nutrient pool size (long-term average of N) in a system with two plant species. Consequences of PMSF on nutrient pool size for every combination of (fL and fN) from (0.0, 0.0) to (1.0, 1.0) with interval (ΔfL, ΔfN) = (0.01, 0.01) are plotted against the average litter decomposability in a system with microbial functional group MR only (red dots), MS only (green dots), and with two competing microbial groups (blue dots). All parameters are set as default values (
, from t = 45,000 to t = 50,000) and the average nutrient pool size (long-term average of N) in a system with two plant species. Consequences of PMSF on nutrient pool size for every combination of (fL and fN) from (0.0, 0.0) to (1.0, 1.0) with interval (ΔfL, ΔfN) = (0.01, 0.01) are plotted against the average litter decomposability in a system with microbial functional group MR only (red dots), MS only (green dots), and with two competing microbial groups (blue dots). All parameters are set as default values (
References
-
- Wedin DA, Tilman D. Species effects on nitrogen cycling: A test with perennial grasses. Oecologia. 1990;84:433–441. - PubMed
-
- Scott NA, Binkley D. Foliage litter quality and annual net N mineralization: comparison across North American forest sites. Oecologia. 1997;111:151–159. - PubMed
-
- Binkley D, Giardina C. Why do tree species affect soils? The warp and woof of tree-soil interactions. Biogeochemistry. 1998;42:89–106.
-
- Berendse F, Oudhof H, Bol J. A comparative study on nutrient cycling in wet heathland ecosystems. I. Litter production and nutrient losses from the plant. Oecologia. 1987;74:174–184. - PubMed
-
- Berendse F, Bobbink R, Rouwenhorst G. A comparative study on nutrient cycling in wet heathland ecosystems. II. Litter decomposition and nutrient mineralization. Oecologia. 1989;78:338–348. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
