

An epigenetic chromatin remodeling role for NFATc1 in transcriptional regulation of growth and survival genes in diffuse large B-cell lymphomas
- PMID: 20664054
- PMCID: PMC2981542
- DOI: 10.1182/blood-2009-12-257378
An epigenetic chromatin remodeling role for NFATc1 in transcriptional regulation of growth and survival genes in diffuse large B-cell lymphomas
Abstract
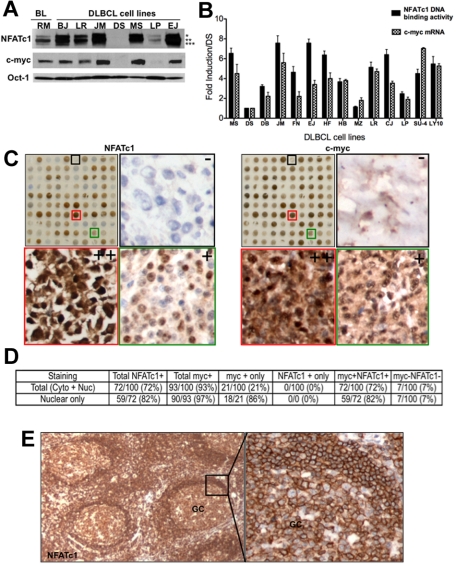
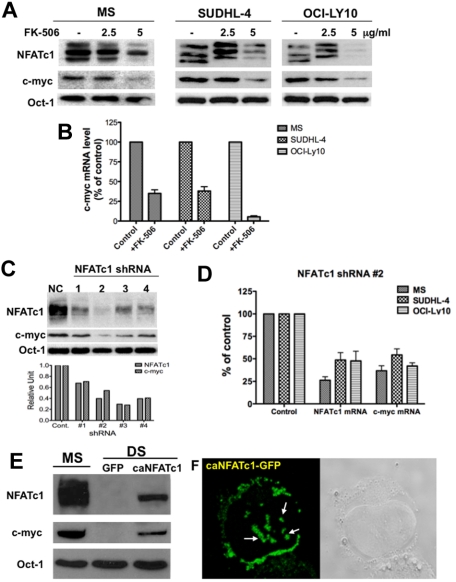
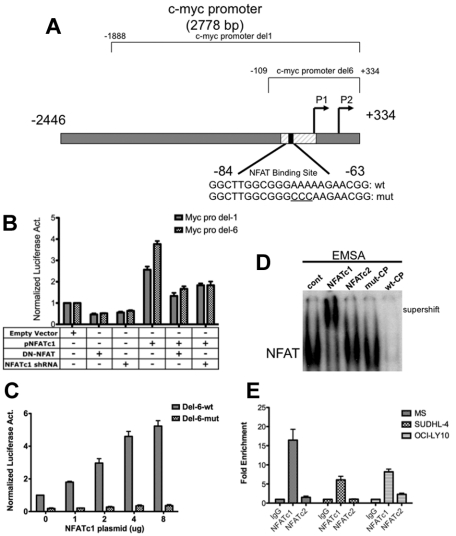
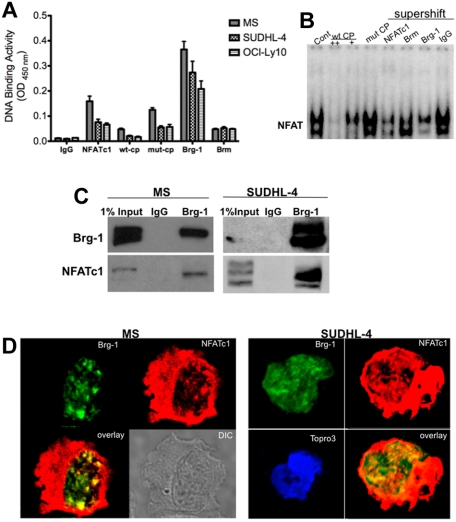
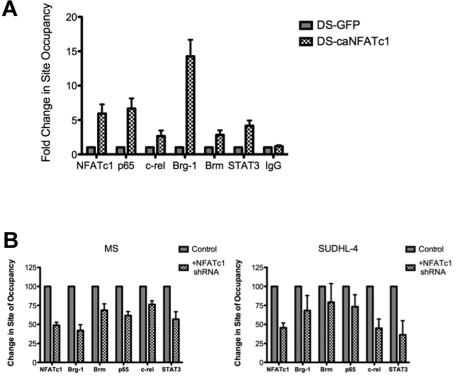
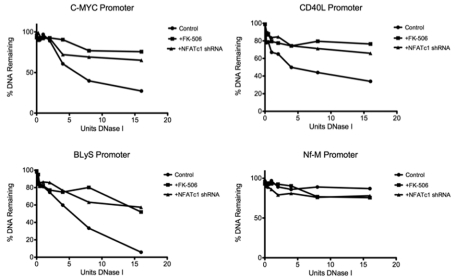
The nuclear factor of activated T cells (NFAT) family of transcription factors functions as integrators of multiple signaling pathways by binding to chromatin in combination with other transcription factors and coactivators to regulate genes central for cell growth and survival in hematopoietic cells. Recent experimental evidence has implicated the calcineurin/NFAT signaling pathway in the pathogenesis of various malignancies, including diffuse large B-cell lymphoma (DLBCL). However, the molecular mechanism(s) underlying NFATc1 regulation of genes controlling lymphoma cell growth and survival is still unclear. In this study, we demonstrate that the transcription factor NFATc1 regulates gene expression in DLBCL cells through a chromatin remodeling mechanism that involves recruitment of the SWItch/Sucrose NonFermentable chromatin remodeling complex ATPase enzyme SMARCA4 (also known as Brahma-related gene 1) to NFATc1 targeted gene promoters. The NFATc1/Brahma-related gene 1 complex induces promoter DNase I hypersensitive sites and recruits other transcription factors to the active chromatin site to regulate gene transcription. Targeting NFATc1 with specific small hairpin RNA inhibits DNase I hypersensitive site formation and down-regulates target gene expression. Our data support a novel epigenetic control mechanism for the transcriptional regulation of growth and survival genes by NFATc1 in the pathophysiology of DLBCL and suggests that targeting NFATc1 could potentially have therapeutic value.
Figures
Similar articles
-
Identification of NFAT binding sites that mediate stimulation of cathepsin K promoter activity by RANK ligand.Gene. 2009 Oct 15;446(2):90-8. doi: 10.1016/j.gene.2009.06.013. Epub 2009 Jun 27. Gene. 2009. PMID: 19563866
-
Constitutive NF-kappaB and NFAT activation in aggressive B-cell lymphomas synergistically activates the CD154 gene and maintains lymphoma cell survival.Blood. 2005 Dec 1;106(12):3940-7. doi: 10.1182/blood-2005-03-1167. Epub 2005 Aug 11. Blood. 2005. PMID: 16099873 Free PMC article.
-
IUGR increases chromatin-remodeling factor Brg1 expression and binding to GR exon 1.7 promoter in newborn male rat hippocampus.Am J Physiol Regul Integr Comp Physiol. 2015 Jul 15;309(2):R119-27. doi: 10.1152/ajpregu.00495.2014. Epub 2015 May 13. Am J Physiol Regul Integr Comp Physiol. 2015. PMID: 25972460
-
Epigenetic regulation in human melanoma: past and future.Epigenetics. 2015;10(2):103-21. doi: 10.1080/15592294.2014.1003746. Epigenetics. 2015. PMID: 25587943 Free PMC article. Review.
-
Architecture and expression of the nfatc1 gene in lymphocytes.Front Immunol. 2014 Feb 3;5:21. doi: 10.3389/fimmu.2014.00021. eCollection 2014. Front Immunol. 2014. PMID: 24550910 Free PMC article. Review.
Cited by
-
Primary CNS lymphoproliferative disease, mycophenolate and calcineurin inhibitor usage.Oncotarget. 2015 Oct 20;6(32):33849-66. doi: 10.18632/oncotarget.5292. Oncotarget. 2015. PMID: 26460822 Free PMC article.
-
Role of NFAT in Chronic Lymphocytic Leukemia and Other B-Cell Malignancies.Front Oncol. 2021 Apr 1;11:651057. doi: 10.3389/fonc.2021.651057. eCollection 2021. Front Oncol. 2021. PMID: 33869054 Free PMC article. Review.
-
Targetable vulnerability of deregulated FOXM1/PLK1 signaling axis in diffuse large B cell lymphoma.Am J Cancer Res. 2022 Oct 15;12(10):4666-4679. eCollection 2022. Am J Cancer Res. 2022. PMID: 36381323 Free PMC article.
-
Zeb1-induced metabolic reprogramming of glycolysis is essential for macrophage polarization in breast cancer.Cell Death Dis. 2022 Mar 4;13(3):206. doi: 10.1038/s41419-022-04632-z. Cell Death Dis. 2022. PMID: 35246504 Free PMC article.
-
NFATC1 promotes cell growth and tumorigenesis in ovarian cancer up-regulating c-Myc through ERK1/2/p38 MAPK signal pathway.Tumour Biol. 2016 Apr;37(4):4493-500. doi: 10.1007/s13277-015-4245-x. Epub 2015 Oct 26. Tumour Biol. 2016. PMID: 26501422
References
-
- Alexander DD, Mink PJ, Adami HO, et al. The non-Hodgkin lymphomas: a review of the epidemiologic literature. Int J Cancer. 2007;(120 suppl 12):1–39. - PubMed
-
- Armitage JO, Weisenburger DD. New approach to classifying non-Hodgkin's lymphomas: clinical features of the major histologic subtypes. Non-Hodgkin's Lymphoma Classification Project. J Clin Oncol. 1998;16(8):2780–2795. - PubMed
-
- Fisher RI, Shah P. Current trends in large cell lymphoma. Leukemia. 2003;17(10):1948–1960. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
