

Essential role for focal adhesion kinase in regulating stress hematopoiesis
- PMID: 20664055
- PMCID: PMC2993618
- DOI: 10.1182/blood-2010-01-262790
Essential role for focal adhesion kinase in regulating stress hematopoiesis
Abstract
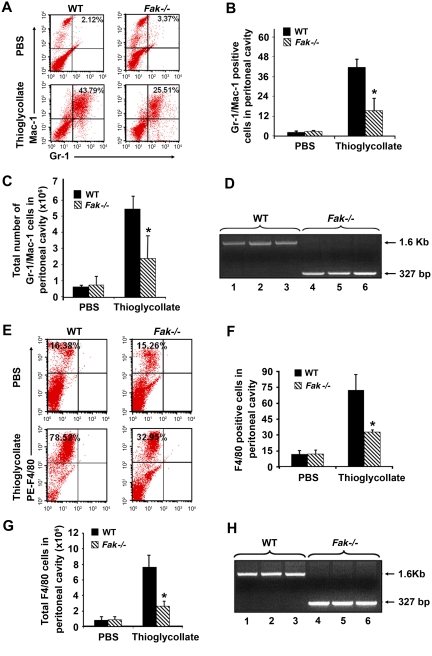
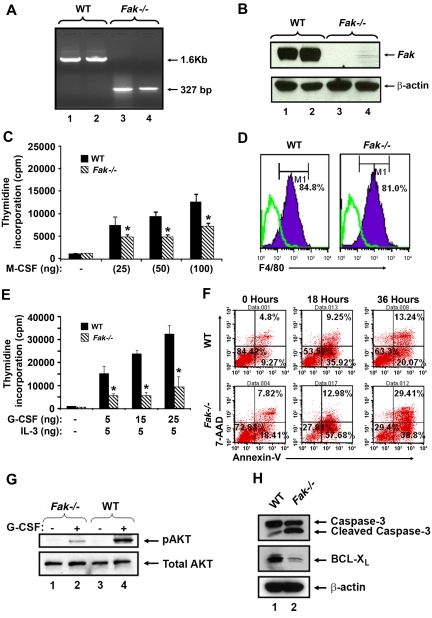
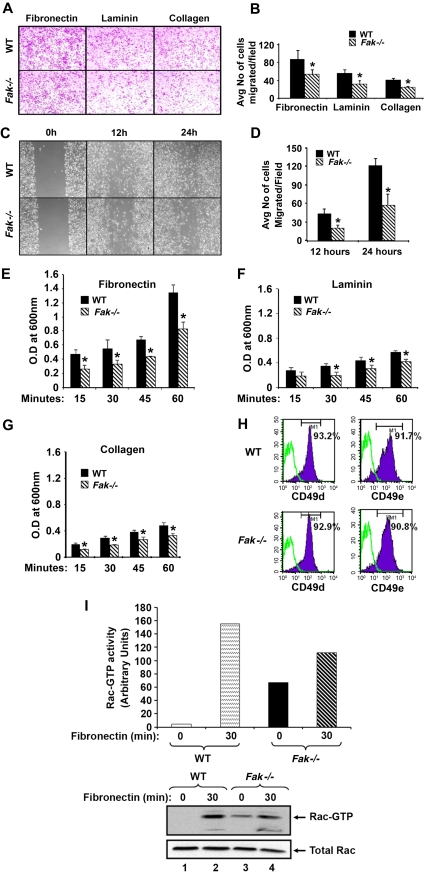
Focal adhesion kinase (FAK) is a non-receptor tyrosine kinase that has been extensively studied in fibroblasts; however its function in hematopoiesis remains an enigma. FAK is thought to be expressed in myeloid and erythroid progenitors, and its expression is enhanced in response to cytokines such as granu-locyte macrophage colony-stimulating factor. Furthermore, bone marrow cells cultured in granulocyte macrophage colony-stimulating factor show active migration and chemoattractant-induced polarization, which correlates with FAK induction. While loss of FAK in mice results in embryonic lethality, we have deleted FAK in the adult bone marrow. We show an essential role for FAK in regulating hemolytic, myelotoxic, as well as acute inflammatory stress responses in vivo. In vitro, loss of FAK in erythroid and myeloid progenitor's results in impaired cytokine induced growth and survival, as well as defects in the activation and expression of antiapoptotic proteins caspase 3 and Bcl-x(L). Additionally, reduced migration and adhesion of myeloid cells on extracellular matrix proteins, as well as impaired activation of Rac GTPase is also observed in the absence of FAK. Our studies reveal an essential role for FAK in integrating growth/survival and adhesion based functions in myeloid and erythroid cells predominantly under conditions of stress.
Figures
Similar articles
-
Focal adhesion kinase upregulated by granulocyte-macrophage colony-stimulating factor but not by interleukin-3 in differentiating myeloid cells.Blood. 1997 May 1;89(9):3434-42. Blood. 1997. PMID: 9129051
-
The immunosuppressant rapamycin blocks in vitro responses to hematopoietic cytokines and inhibits recovering but not steady-state hematopoiesis in vivo.Blood. 1994 Sep 1;84(5):1543-52. Blood. 1994. PMID: 7520778
-
FAK Deficiency in Bone Marrow Stromal Cells Alters Their Homeostasis and Drives Abnormal Proliferation and Differentiation of Haematopoietic Stem Cells.Cells. 2020 Mar 6;9(3):646. doi: 10.3390/cells9030646. Cells. 2020. PMID: 32155953 Free PMC article.
-
Focal adhesion kinase: from in vitro studies to functional analyses in vivo.Curr Protein Pept Sci. 2011 Feb;12(1):52-67. doi: 10.2174/138920311795659452. Curr Protein Pept Sci. 2011. PMID: 21190526 Review.
-
Focal adhesion kinase controls prostate cancer progression via intrinsic kinase and scaffolding functions.Anticancer Agents Med Chem. 2011 Sep;11(7):607-16. doi: 10.2174/187152011796817646. Anticancer Agents Med Chem. 2011. PMID: 21355844 Review.
Cited by
-
Fak depletion in both hematopoietic and nonhematopoietic niche cells leads to hematopoietic stem cell expansion.Exp Hematol. 2012 Apr;40(4):307-17.e3. doi: 10.1016/j.exphem.2011.11.010. Epub 2011 Dec 8. Exp Hematol. 2012. PMID: 22155722 Free PMC article.
-
FAK mediates a compensatory survival signal parallel to PI3K-AKT in PTEN-null T-ALL cells.Cell Rep. 2015 Mar 31;10(12):2055-68. doi: 10.1016/j.celrep.2015.02.056. Epub 2015 Mar 19. Cell Rep. 2015. PMID: 25801032 Free PMC article.
-
SWAP-70 regulates erythropoiesis by controlling α4 integrin.Haematologica. 2011 Dec;96(12):1743-52. doi: 10.3324/haematol.2011.050468. Epub 2011 Aug 31. Haematologica. 2011. PMID: 21880631 Free PMC article.
-
Rho kinase regulates the survival and transformation of cells bearing oncogenic forms of KIT, FLT3, and BCR-ABL.Cancer Cell. 2011 Sep 13;20(3):357-69. doi: 10.1016/j.ccr.2011.07.016. Cancer Cell. 2011. PMID: 21907926 Free PMC article.
-
ROCK1 functions as a critical regulator of stress erythropoiesis and survival by regulating p53.Blood. 2012 Oct 4;120(14):2868-78. doi: 10.1182/blood-2011-10-384172. Epub 2012 Aug 13. Blood. 2012. PMID: 22889758 Free PMC article.
References
-
- Guan JL, Shalloway D. Regulation of focal adhesion-associated protein tyrosine kinase by both cellular adhesion and oncogenic transformation. Nature. 1992;358(6388):690–692. - PubMed
-
- Parsons JT. Focal adhesion kinase: the first ten years. J Cell Sci. 2003;116(Pt 8):1409–1416. - PubMed
-
- Sieg DJ, Hauck CR, Ilic D, et al. FAK integrates growth-factor and integrin signals to promote cell migration. Nat Cell Biol. 2000;2(5):249–256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
