

Muscle KATP channels: recent insights to energy sensing and myoprotection
- PMID: 20664073
- PMCID: PMC3125986
- DOI: 10.1152/physrev.00027.2009
Muscle KATP channels: recent insights to energy sensing and myoprotection
Abstract
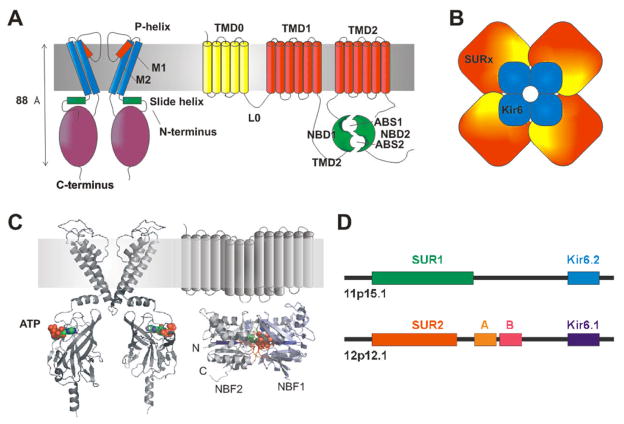
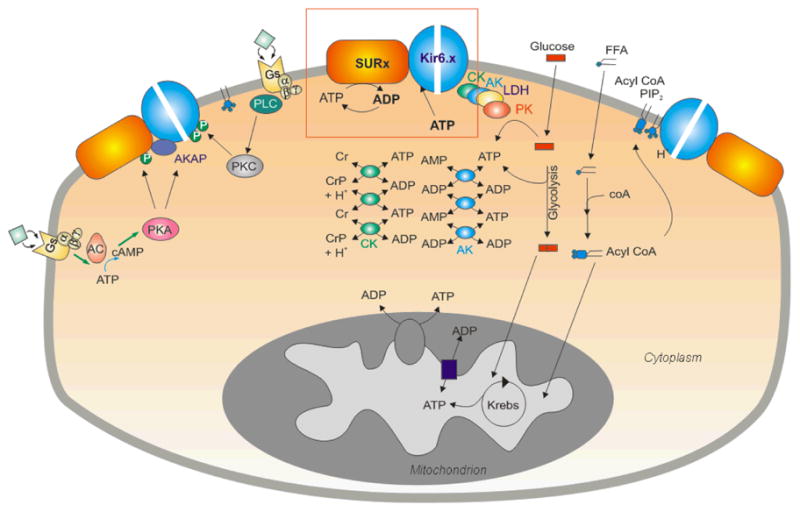
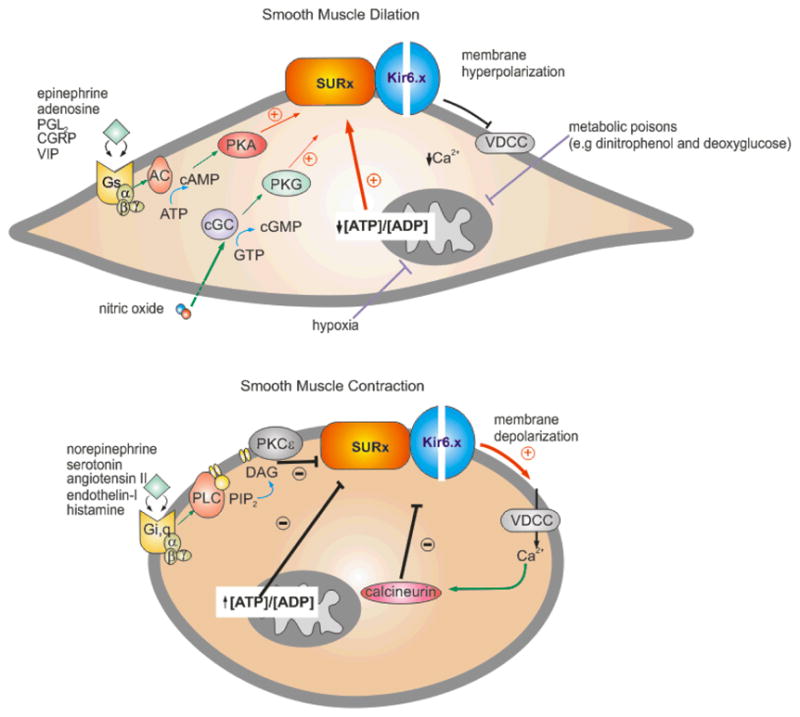
ATP-sensitive potassium (K(ATP)) channels are present in the surface and internal membranes of cardiac, skeletal, and smooth muscle cells and provide a unique feedback between muscle cell metabolism and electrical activity. In so doing, they can play an important role in the control of contractility, particularly when cellular energetics are compromised, protecting the tissue against calcium overload and fiber damage, but the cost of this protection may be enhanced arrhythmic activity. Generated as complexes of Kir6.1 or Kir6.2 pore-forming subunits with regulatory sulfonylurea receptor subunits, SUR1 or SUR2, the differential assembly of K(ATP) channels in different tissues gives rise to tissue-specific physiological and pharmacological regulation, and hence to the tissue-specific pharmacological control of contractility. The last 10 years have provided insights into the regulation and role of muscle K(ATP) channels, in large part driven by studies of mice in which the protein determinants of channel activity have been deleted or modified. As yet, few human diseases have been correlated with altered muscle K(ATP) activity, but genetically modified animals give important insights to likely pathological roles of aberrant channel activity in different muscle types.
Figures
References
-
- Aguilar-Bryan L, Nichols CG, Rajan AS, Parker C, Bryan J. Co-expression of sulfonylurea receptors and KATP channels in hamster insulinoma tumor (HIT) cells. Evidence for direct association of the receptor with the channel. J Biol Chem. 1992;267:14934–14940. - PubMed
-
- Aguilar-Bryan L, Nichols CG, Wechsler SW, Clement JPt, Boyd AE, 3rd, Gonzalez G, Herrera-Sosa H, Nguy K, Bryan J, Nelson DA. Cloning of the beta cell high-affinity sulfonylurea receptor: a regulator of insulin secretion. Science. 1995;268:423–426. - PubMed
-
- Allard B, Lazdunski M. Nucleotide diphosphates activate the ATP-sensitive potassium channel in mouse skeletal muscle. Pflugers Arch. 1992;422:185–192. - PubMed
-
- Allard B, Lazdunski M. Pharmacological properties of ATP-sensitive K+ channels in mammalian skeletal muscle cells. Eur J Pharmacol. 1993;236:419–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources