

Sema3E-Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice
- PMID: 20664171
- PMCID: PMC2912191
- DOI: 10.1172/JCI42118
Sema3E-Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice
Erratum in
- J Clin Invest. 2011 Jul 1;121(7):2945. Mazzone, Massimilano [corrected to Mazzone, Massimiliano]
Abstract
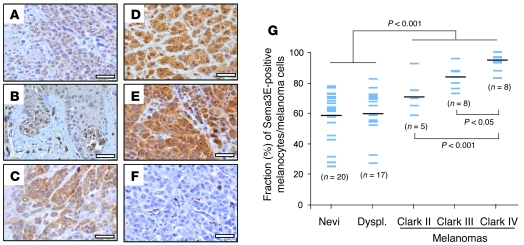
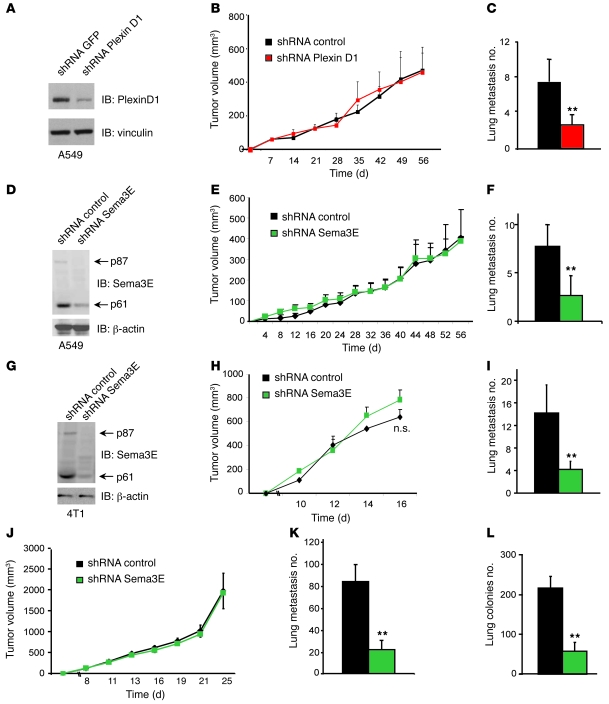
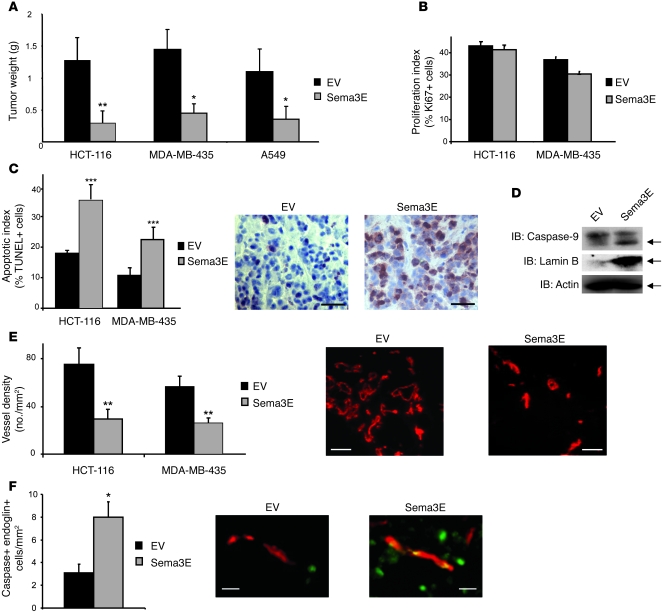
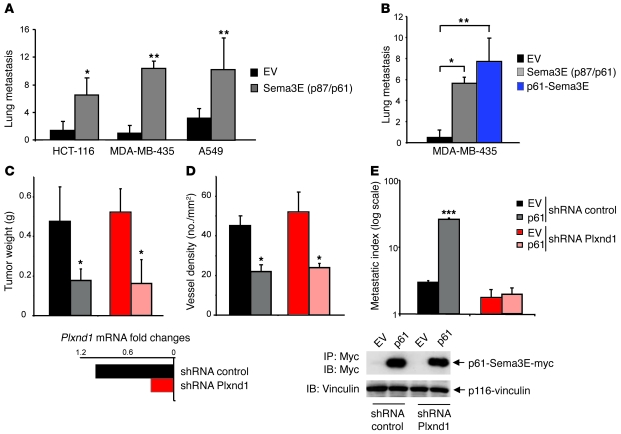
Semaphorin 3E (Sema3E) is a secreted molecule implicated in axonal path finding and inhibition of developmental and postischemic angiogenesis. Sema3E is also highly expressed in metastatic cancer cells, but its mechanistic role in tumor progression was not understood. Here we show that expression of Sema3E and its receptor Plexin D1 correlates with the metastatic progression of human tumors. Consistent with the clinical data, knocking down endogenous expression of either Sema3E or Plexin D1 in human metastatic carcinoma cells hampered their metastatic potential when injected into mice, while tumor growth was not markedly affected. Conversely, overexpression of exogenous Sema3E in cancer cells increased their invasiveness, transendothelial migration, and metastatic spreading, although it inhibited tumor vessel formation, resulting in reduced tumor growth in mice. The proinvasive and metastatic activity of Sema3E in tumor cells was dependent on transactivation of the Plexin D1-associated ErbB2/Neu oncogenic kinase. In sum, Sema3E-Plexin D1 signaling in cancer cells is crucially implicated in their metastatic behavior and may therefore be a promising target for strategies aimed at blocking tumor metastasis.
Figures
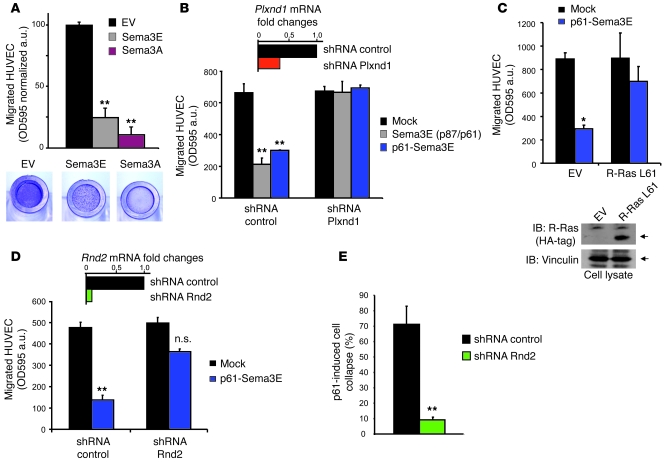
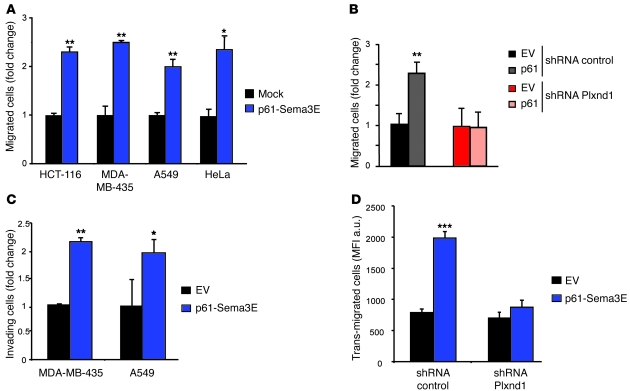
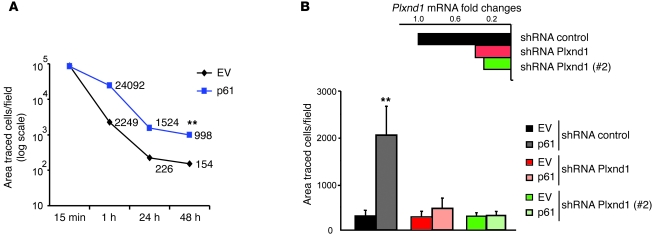
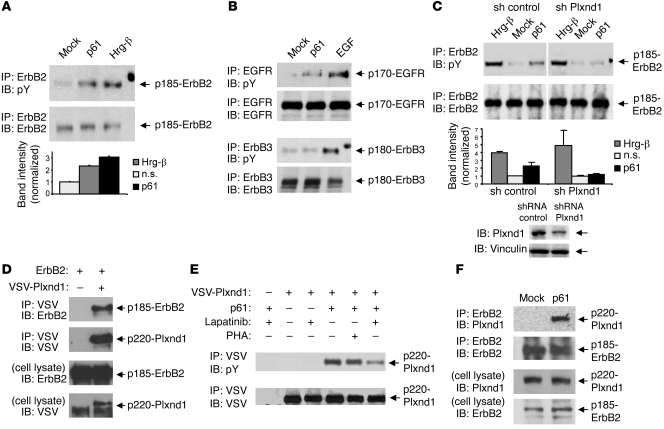
Comment in
-
Semaphorin 3E, an exception to the rule.J Clin Invest. 2010 Aug;120(8):2658-60. doi: 10.1172/JCI44110. Epub 2010 Jul 26. J Clin Invest. 2010. PMID: 20664165 Free PMC article.
References
-
- Casazza A, Fazzari P, Tamagnone L. Semaphorin signals in cell adhesion and cell migration: functional role and molecular mechanisms. Adv Exp Med Biol. 2007;600:90–108. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
