Competitive interactions in Escherichia coli populations: the role of bacteriocins
- PMID: 20664553
- PMCID: PMC3105674
- DOI: 10.1038/ismej.2010.90
Competitive interactions in Escherichia coli populations: the role of bacteriocins
Abstract
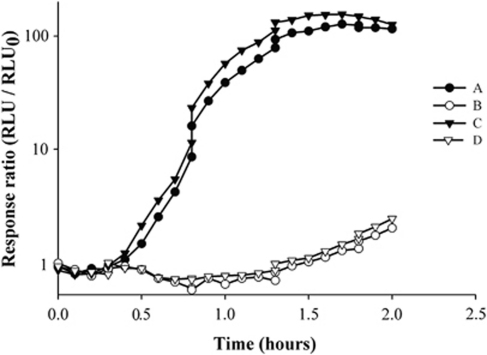
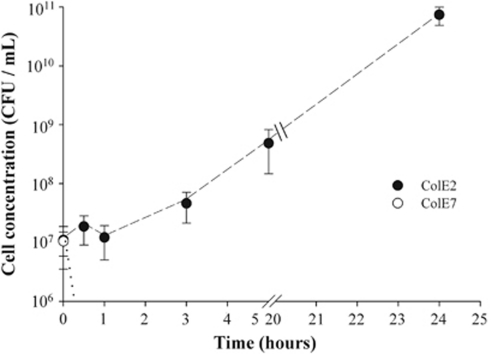
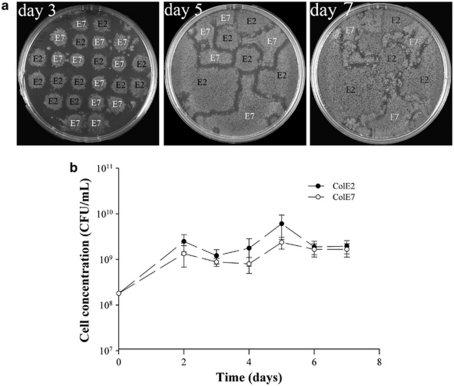
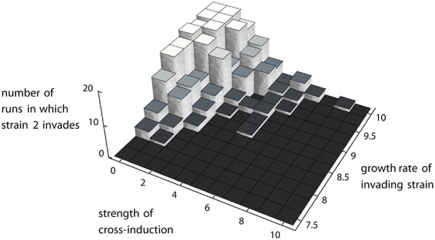
Explaining the coexistence of competing species is a major challenge in community ecology. In bacterial systems, competition is often driven by the production of bacteriocins, which are narrow-spectrum proteinaceous toxins that serve to kill closely related species, providing the producer better access to limited resources. Bacteriocin producers have been shown to competitively exclude sensitive, nonproducing strains. However, the dynamics between bacteriocin producers, each lethal to its competitor, are largely unknown. In this study, we used in vitro, in vivo and in silico models to study competitive interactions between bacteriocin producers. Two Escherichia coli strains were generated, each carrying a DNA-degrading bacteriocin (colicins E2 and E7). Using reporter-gene assays, we showed that each DNase bacteriocin is not only lethal to its opponent but, at lower doses, can also induce the expression of its opponent's toxin. In a well-mixed habitat, the E2 producer outcompeted its adversary; however, in structured environments (on plates or in mice colons), the two producers coexisted in a spatially 'frozen' pattern. Coexistence occurred when the producers were initiated with a clumped spatial distribution. This suggests that a 'clump' of each producer can block invasion of the other producer. Agent-based simulation of bacteriocin-mediated competition further showed that mutual exclusion in a structured environment is a relatively robust result. These models imply that colicin-mediated colicin induction enables producers to successfully compete and defend their niche against invaders. This suggests that localized interactions between producers of DNA-degrading toxins can lead to stable coexistence of heterogeneously distributed strains within the bacterial community and to the maintenance of diversity.
Figures