

Intragenomic conflict in populations infected by Parthenogenesis Inducing Wolbachia ends with irreversible loss of sexual reproduction
- PMID: 20667099
- PMCID: PMC2927591
- DOI: 10.1186/1471-2148-10-229
Intragenomic conflict in populations infected by Parthenogenesis Inducing Wolbachia ends with irreversible loss of sexual reproduction
Abstract
Background: The maternally inherited, bacterial symbiont, parthenogenesis inducing (PI) Wolbachia, causes females in some haplodiploid insects to produce daughters from both fertilized and unfertilized eggs. The symbionts, with their maternal inheritance, benefit from inducing the production of exclusively daughters, however the optimal sex ratio for the nuclear genome is more male-biased. Here we examine through models how an infection with PI-Wolbachia in a previously uninfected population leads to a genomic conflict between PI-Wolbachia and the nuclear genome. In most natural populations infected with PI-Wolbachia the infection has gone to fixation and sexual reproduction is impossible, specifically because the females have lost their ability to fertilize eggs, even when mated with functional males.
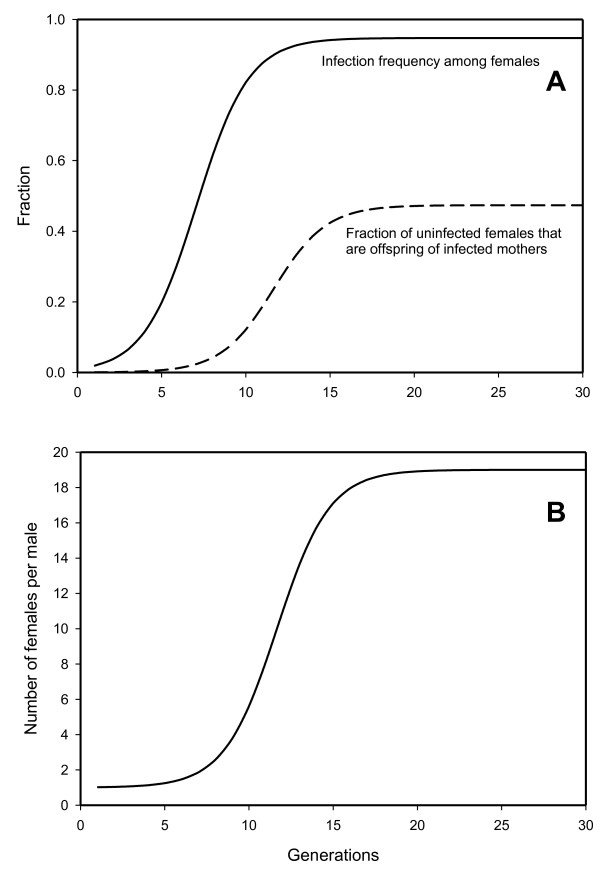
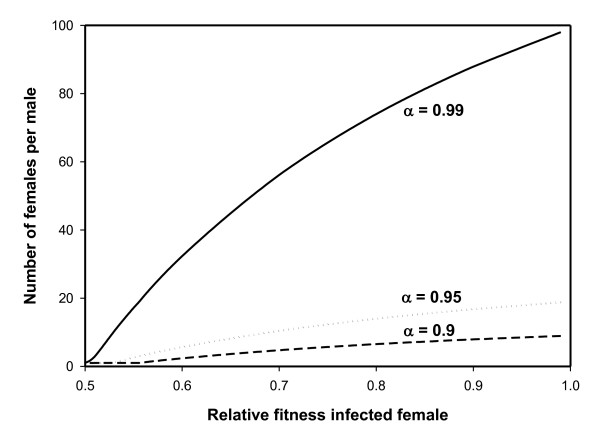
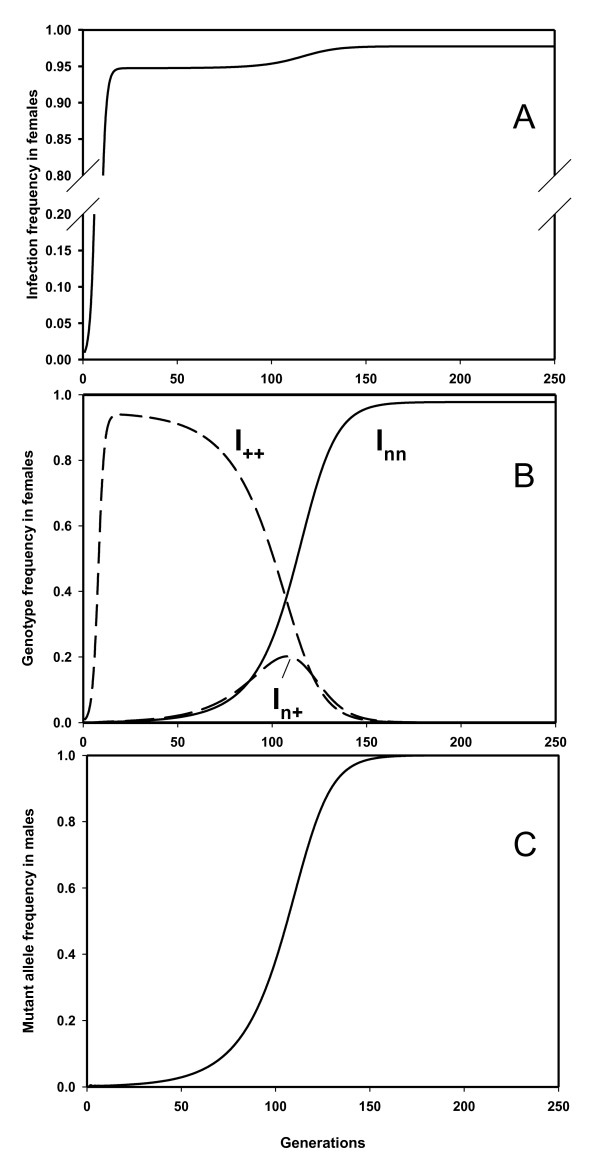
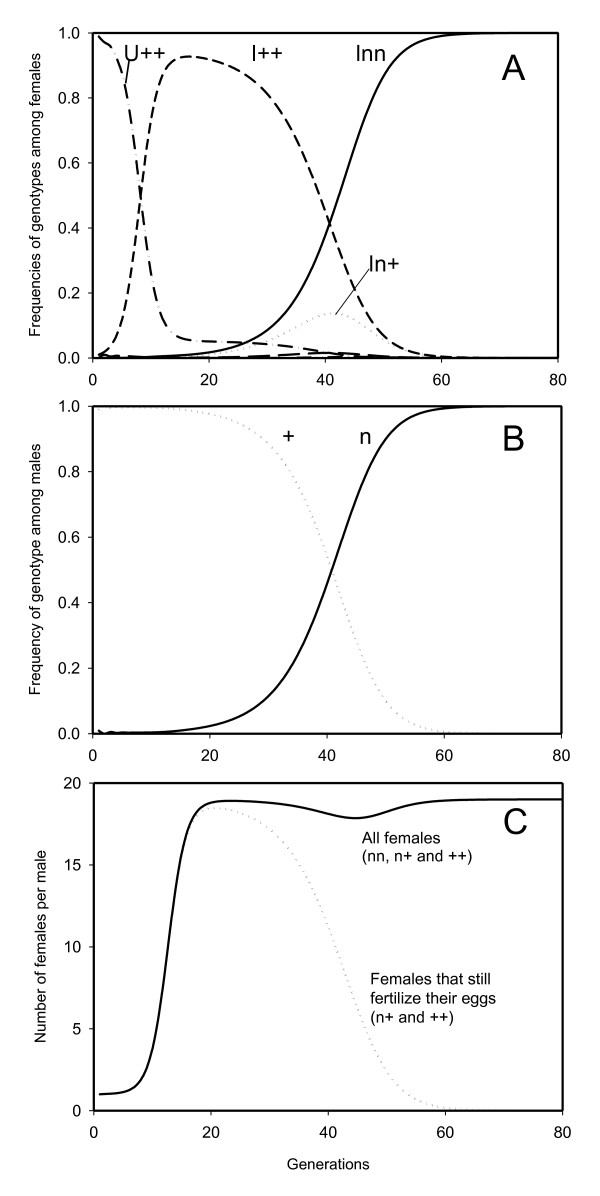
Results: The PI Wolbachia infection by itself does not interfere with the fertilization process in infected eggs, fertilized infected eggs develop into biparental infected females. Because of the increasingly female-biased sex ratio in the population during a spreading PI-Wolbachia infection, sex allocation alleles in the host that cause the production of more sons are rapidly selected. In haplodiploid species a reduced fertilization rate leads to the production of more sons. Selection for the reduced fertilization rate leads to a spread of these alleles through both the infected and uninfected population, eventually resulting in the population becoming fixed for both the PI-Wolbachia infection and the reduced fertilization rate. Fertilization rate alleles that completely interfere with fertilization ("virginity alleles") will be selected over alleles that still allow for some fertilization. This drives the final resolution of the conflict: the irreversible loss of sexual reproduction and the complete dependence of the host on its symbiont.
Conclusions: This study shows that dependence among organisms can evolve rapidly due to the resolution of the conflicts between cytoplasmic and nuclear genes, and without requiring a mutualism between the partners.
Figures
Similar articles
-
Genetics of female functional virginity in the parthenogenesis-Wolbachia infected parasitoid wasp Telenomus nawai (Hymenoptera: Scelionidae).Heredity (Edinb). 2005 Apr;94(4):402-7. doi: 10.1038/sj.hdy.6800617. Heredity (Edinb). 2005. PMID: 15523503
-
The genetics and evolution of obligate reproductive parasitism in Trichogramma pretiosum infected with parthenogenesis-inducing Wolbachia.Heredity (Edinb). 2011 Jan;106(1):58-67. doi: 10.1038/hdy.2010.48. Epub 2010 May 5. Heredity (Edinb). 2011. PMID: 20442735 Free PMC article.
-
The effects of outbreeding on a parasitoid wasp fixed for infection with a parthenogenesis-inducing Wolbachia symbiont.Heredity (Edinb). 2017 Dec;119(6):411-417. doi: 10.1038/hdy.2017.53. Epub 2017 Sep 13. Heredity (Edinb). 2017. PMID: 28902190 Free PMC article.
-
Reproductive parasitism: maternally inherited symbionts in a biparental world.Cold Spring Harb Perspect Biol. 2015 May 1;7(5):a017699. doi: 10.1101/cshperspect.a017699. Cold Spring Harb Perspect Biol. 2015. PMID: 25934011 Free PMC article. Review.
-
Wolbachia pipientis: microbial manipulator of arthropod reproduction.Annu Rev Microbiol. 1999;53:71-102. doi: 10.1146/annurev.micro.53.1.71. Annu Rev Microbiol. 1999. PMID: 10547686 Review.
Cited by
-
Living in the endosymbiotic world of Wolbachia: A centennial review.Cell Host Microbe. 2021 Jun 9;29(6):879-893. doi: 10.1016/j.chom.2021.03.006. Epub 2021 May 3. Cell Host Microbe. 2021. PMID: 33945798 Free PMC article. Review.
-
Losing the desire: selection can promote obligate asexuality.BMC Biol. 2010 Jul 28;8:101. doi: 10.1186/1741-7007-8-101. BMC Biol. 2010. PMID: 20687906 Free PMC article.
-
A single QTL with large effect is associated with female functional virginity in an asexual parasitoid wasp.Mol Ecol. 2021 May;30(9):1979-1992. doi: 10.1111/mec.15863. Epub 2021 Mar 15. Mol Ecol. 2021. PMID: 33638236 Free PMC article.
-
Host and symbiont genetic contributions to fitness in a Trichogramma-Wolbachia symbiosis.PeerJ. 2018 Apr 19;6:e4655. doi: 10.7717/peerj.4655. eCollection 2018. PeerJ. 2018. PMID: 29686946 Free PMC article.
-
Distinct epigenomic and transcriptomic modifications associated with Wolbachia-mediated asexuality.PLoS Pathog. 2020 Mar 18;16(3):e1008397. doi: 10.1371/journal.ppat.1008397. eCollection 2020 Mar. PLoS Pathog. 2020. PMID: 32187233 Free PMC article.
References
-
- Cosmides LM, Tooby J. Cytoplasmatic inheritance and intragenomic conflict. JtheorBiol. 1981;89:83–129. - PubMed
-
- Hurst LD. Intragenomic conflict as an evolutionary force. Proc R Soc Lond. 1992;248:135–140. doi: 10.1098/rspb.1992.0053. - DOI
-
- Eberhard WG. Evolutionary consequences of intracellular organelle competition. QuartRevBiol. 1980;55:231–249. - PubMed
-
- Hoekstra RF. In: Experientia Supplementum (Basel), Vol 55 The Evolution of Sex and Its Consequences 403p Birkhaeuser. Stearns SC, editor. Verlag: Basel, Switzerland; Boston, Massachusetts, USA Illus; 1987. The Evolution of Sexes; pp. 59–92.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
