

MMP13 mediates cell cycle progression in melanocytes and melanoma cells: in vitro studies of migration and proliferation
- PMID: 20667128
- PMCID: PMC2915980
- DOI: 10.1186/1476-4598-9-201
MMP13 mediates cell cycle progression in melanocytes and melanoma cells: in vitro studies of migration and proliferation
Abstract
Background: Melanoma cells are usually characterized by a strong proliferative potential and efficient invasive migration. Among the multiple molecular changes that are recorded during progression of this disease, aberrant activation of receptor tyrosine kinases (RTK) is often observed. Activation of matrix metalloproteases goes along with RTK activation and usually enhances RTK-driven migration. The purpose of this study was to examine RTK-driven three-dimensional migration of melanocytes and the pro-tumorigenic role of matrix metalloproteases for melanocytes and melanoma cells.
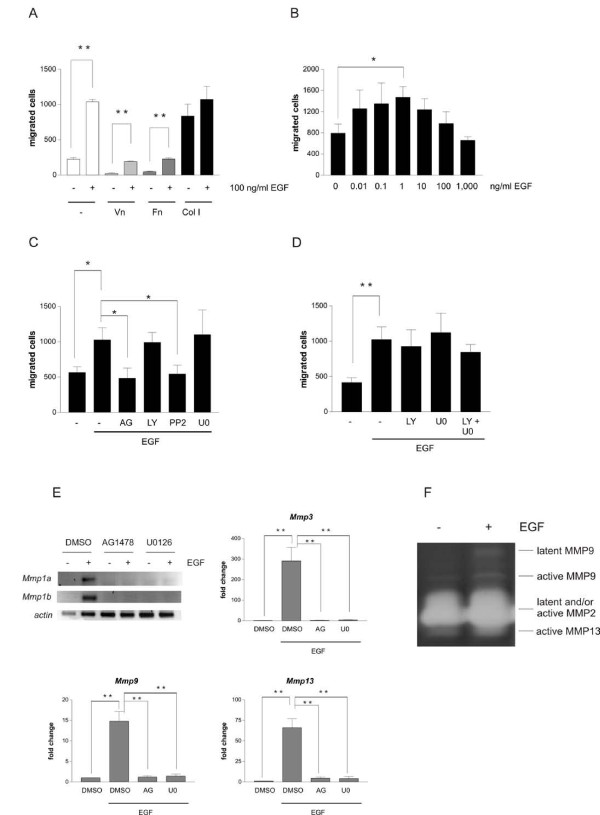
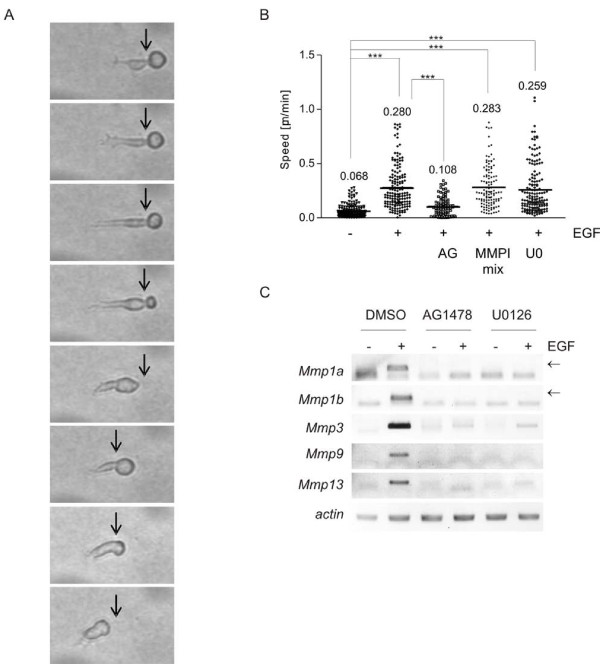
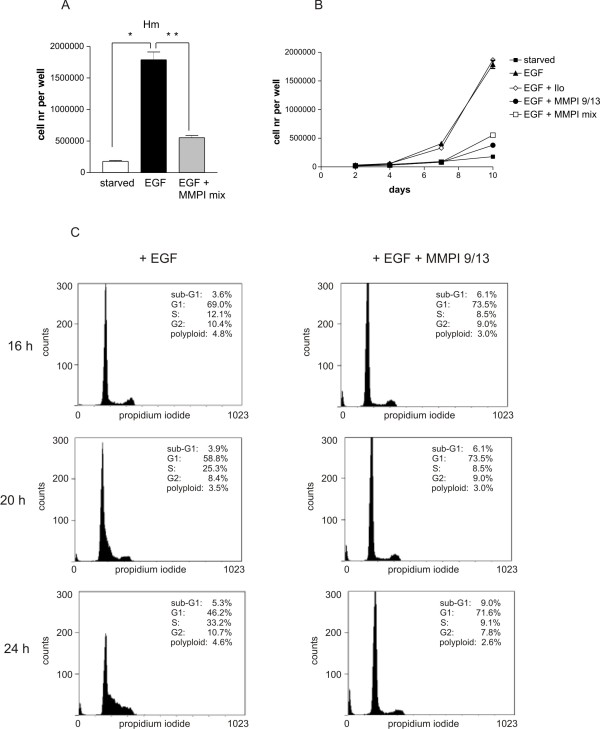
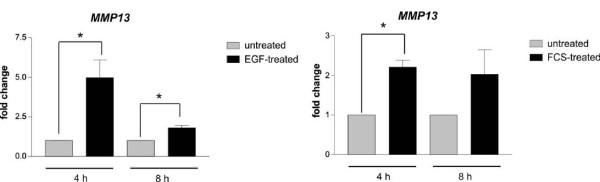
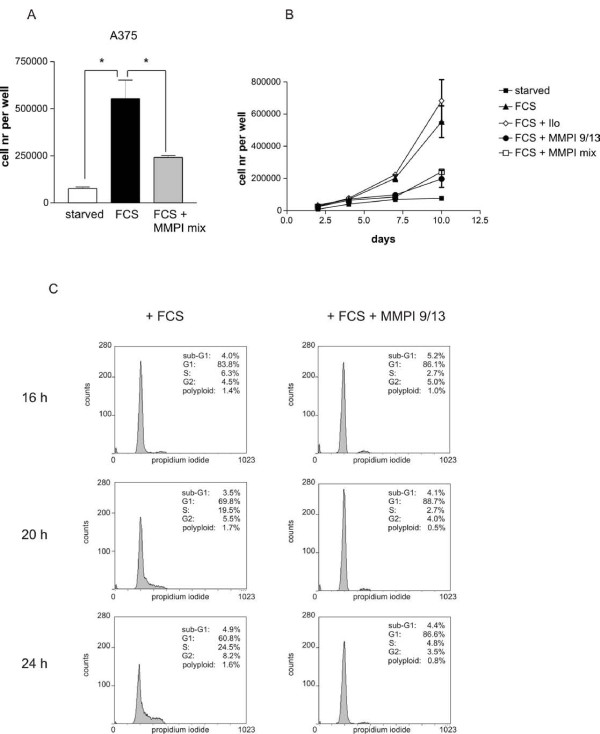
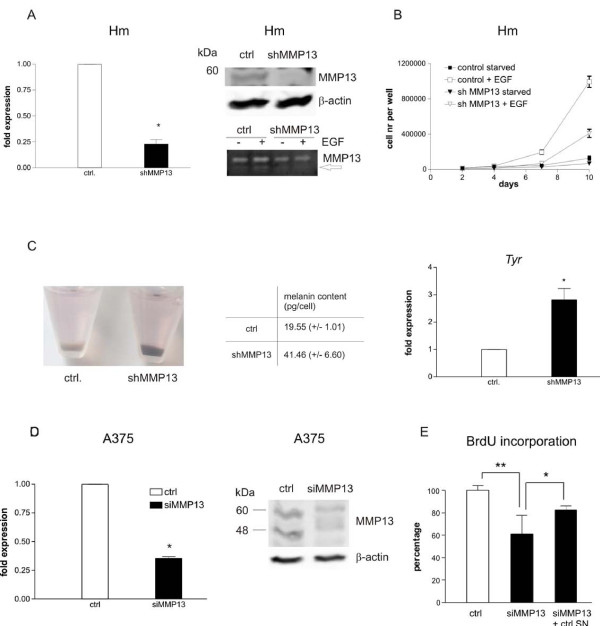
Results: Using experimental melanocyte dedifferentiation as a model for early melanomagenesis we show that an activated EGF receptor variant potentiates migration through three-dimensional fibrillar collagen. EGFR stimulation also resulted in a strong induction of matrix metalloproteases in a MAPK-dependent manner. However, neither MAPK nor MMP activity were required for migration, as the cells migrated in an entirely amoeboid mode. Instead, MMPs fulfilled a function in cell cycle regulation, as their inhibition resulted in strong growth inhibition of melanocytes. The same effect was observed in the human melanoma cell line A375 after stimulation with FCS. Using sh- and siRNA techniques, we could show that MMP13 is the protease responsible for this effect. Along with decreased proliferation, knockdown of MMP13 strongly enhanced pigmentation of melanocytes.
Conclusions: Our data show for the first time that growth stimuli are mediated via MMP13 in melanocytes and melanoma, suggesting an autocrine MMP13-driven loop. Given that MMP13-specific inhibitors are already developed, these results support the evaluation of these inhibitors in the treatment of melanoma.
Figures
Similar articles
-
Gene expression analysis after receptor tyrosine kinase activation reveals new potential melanoma proteins.BMC Cancer. 2010 Jul 21;10:386. doi: 10.1186/1471-2407-10-386. BMC Cancer. 2010. PMID: 20663135 Free PMC article.
-
ErbB receptors mediate both migratory and proliferative activities in human melanocytes and melanoma cells.Melanoma Res. 2005 Feb;15(1):21-8. doi: 10.1097/00008390-200502000-00005. Melanoma Res. 2005. PMID: 15714117
-
The neural guidance receptor Plexin C1 delays melanoma progression.Oncogene. 2013 Oct 10;32(41):4941-9. doi: 10.1038/onc.2012.511. Epub 2012 Nov 19. Oncogene. 2013. PMID: 23160370 Free PMC article.
-
[The Importance of MITF Signaling Pathway in the Regulation of Proliferation and Invasiveness of Malignant Melanoma].Klin Onkol. 2016 Fall;29(5):347-350. doi: 10.14735/amko2016347. Klin Onkol. 2016. PMID: 27739313 Review. Czech.
-
Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells.Pigment Cell Melanoma Res. 2013 May;26(3):316-25. doi: 10.1111/pcmr.12080. Epub 2013 Mar 27. Pigment Cell Melanoma Res. 2013. PMID: 23433358 Review.
Cited by
-
Xmrks the Spot: Fish Models for Investigating Epidermal Growth Factor Receptor Signaling in Cancer Research.Cells. 2021 May 7;10(5):1132. doi: 10.3390/cells10051132. Cells. 2021. PMID: 34067095 Free PMC article. Review.
-
Targets for Renal Carcinoma Growth Control Identified by Screening FOXD1 Cell Proliferation Pathways.Cancers (Basel). 2022 Aug 16;14(16):3958. doi: 10.3390/cancers14163958. Cancers (Basel). 2022. PMID: 36010951 Free PMC article.
-
Inhibition of MMP14 potentiates the therapeutic effect of temozolomide and radiation in gliomas.Cancer Med. 2013 Aug;2(4):457-67. doi: 10.1002/cam4.104. Epub 2013 Jun 30. Cancer Med. 2013. PMID: 24156018 Free PMC article.
-
The BRAF(V600E) inhibitor, PLX4032, increases type I collagen synthesis in melanoma cells.Matrix Biol. 2015 Oct;48:66-77. doi: 10.1016/j.matbio.2015.05.007. Epub 2015 May 16. Matrix Biol. 2015. PMID: 25989506 Free PMC article.
-
Constitutive activation of EGFR is associated with tumor progression and plays a prominent role in malignant phenotype of chondrosarcoma.Oncotarget. 2019 May 7;10(34):3166-3182. doi: 10.18632/oncotarget.26899. eCollection 2019 May 7. Oncotarget. 2019. PMID: 31139331 Free PMC article.
References
-
- Ramnarain DB, Park S, Lee DY, Hatanpaa KJ, Scoggin SO, Otu H, Libermann TA, Raisanen JM, Ashfaq R, Wong ET, Wu J, Elliott R, Habib AA. Differential gene expression analysis reveals generation of an autocrine loop by a mutant epidermal growth factor receptor in glioma cells. Cancer Res. 2006;66:867–874. doi: 10.1158/0008-5472.CAN-05-2753. - DOI - PubMed
-
- Tateishi M, Ishida T, Mitsudomi T, Kaneko S, Sugimachi K. Immunohistochemical evidence of autocrine growth factors in adenocarcinoma of the human lung. Cancer Res. 1990;50:7077–7080. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
