

The tomato odorless-2 mutant is defective in trichome-based production of diverse specialized metabolites and broad-spectrum resistance to insect herbivores
- PMID: 20668059
- PMCID: PMC2938144
- DOI: 10.1104/pp.110.160192
The tomato odorless-2 mutant is defective in trichome-based production of diverse specialized metabolites and broad-spectrum resistance to insect herbivores
Abstract
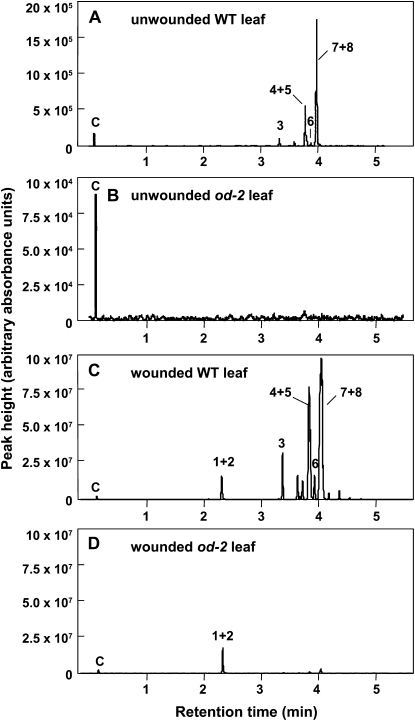
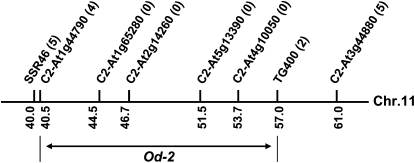
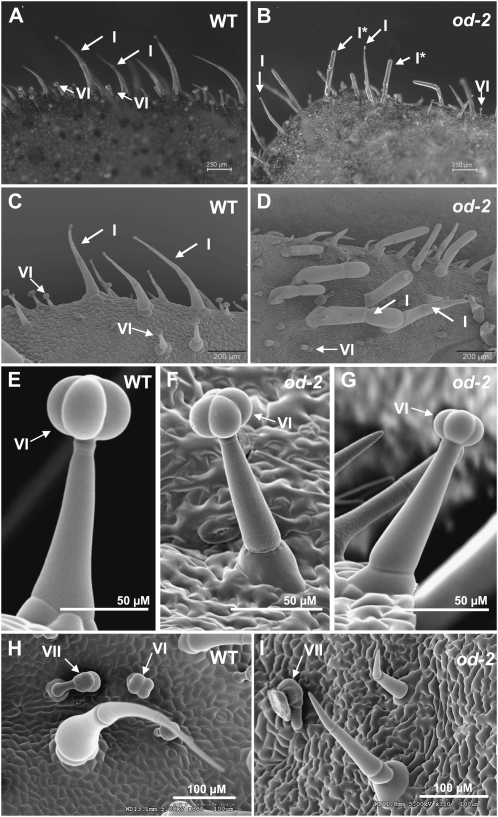
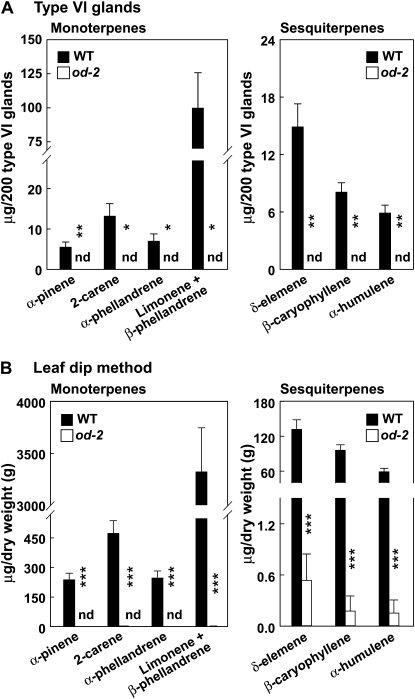
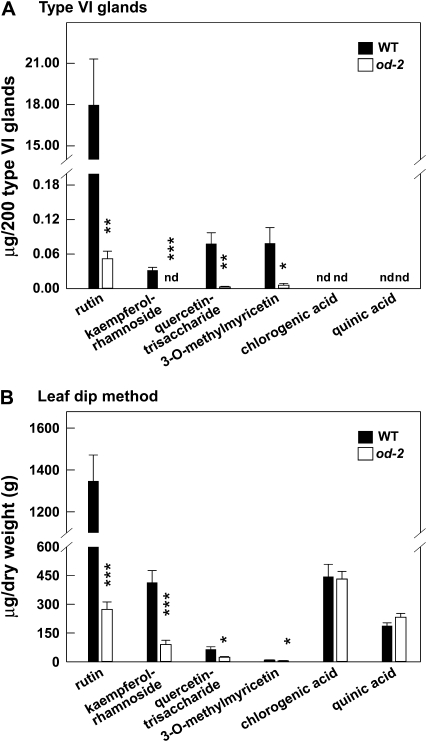
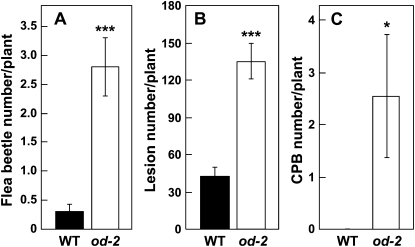
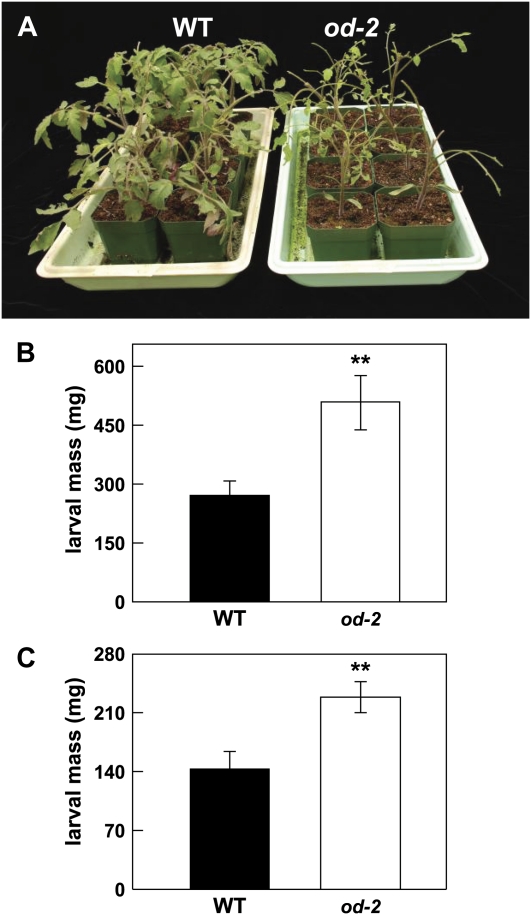
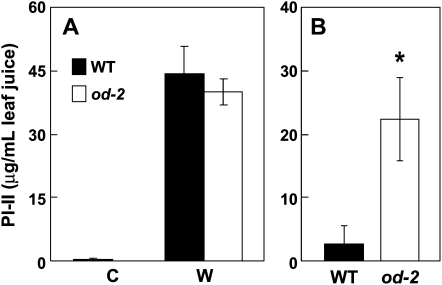
Glandular secreting trichomes of cultivated tomato (Solanum lycopersicum) produce a wide array of volatile and nonvolatile specialized metabolites. Many of these compounds contribute to the characteristic aroma of tomato foliage and constitute a key part of the language by which plants communicate with other organisms in natural environments. Here, we describe a novel recessive mutation called odorless-2 (od-2) that was identified on the basis of an altered leaf-aroma phenotype. od-2 plants exhibit pleiotrophic phenotypes, including alterations in the morphology, density, and chemical composition of glandular trichomes. Type VI glandular trichomes isolated from od-2 leaves accumulate only trace levels of monoterpenes, sesquiterpenes, and flavonoids. Other foliar defensive compounds, including acyl sugars, glycoalkaloids, and jasmonate-regulated proteinase inhibitors, are produced in od-2 leaves. Growth of od-2 plants under natural field conditions showed that the mutant is highly susceptible to attack by an indigenous flea beetle, Epitrix cucumeris, and the Colorado potato beetle, Leptinotarsa decemlineata. The increased susceptibility of od-2 plants to Colorado potato beetle larvae and to the solanaceous specialist Manduca sexta was verified in no-choice bioassays. These findings indicate that Od-2 is essential for the synthesis of diverse trichome-borne compounds and further suggest that these compounds influence host plant selection and herbivore community composition under natural conditions.
Figures

Similar articles
-
Role of trichomes in defense against herbivores: comparison of herbivore response to woolly and hairless trichome mutants in tomato (Solanum lycopersicum).Planta. 2012 Oct;236(4):1053-66. doi: 10.1007/s00425-012-1651-9. Epub 2012 May 3. Planta. 2012. PMID: 22552638
-
Distortion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry.J Exp Bot. 2010 Feb;61(4):1053-64. doi: 10.1093/jxb/erp370. Epub 2009 Dec 16. J Exp Bot. 2010. PMID: 20018901 Free PMC article.
-
Light Intensity-Mediated Induction of Trichome-Associated Allelochemicals Increases Resistance Against Thrips in Tomato.Plant Cell Physiol. 2018 Dec 1;59(12):2462-2475. doi: 10.1093/pcp/pcy166. Plant Cell Physiol. 2018. PMID: 30124946 Free PMC article.
-
Glandular trichomes: micro-organs with model status?New Phytol. 2020 Mar;225(6):2251-2266. doi: 10.1111/nph.16283. Epub 2019 Dec 10. New Phytol. 2020. PMID: 31651036 Review.
-
Genetic Control of Glandular Trichome Development.Trends Plant Sci. 2020 May;25(5):477-487. doi: 10.1016/j.tplants.2019.12.025. Epub 2020 Jan 23. Trends Plant Sci. 2020. PMID: 31983619 Review.
Cited by
-
Exploring the Relationship between Trichome and Terpene Chemistry in Chrysanthemum.Plants (Basel). 2022 May 26;11(11):1410. doi: 10.3390/plants11111410. Plants (Basel). 2022. PMID: 35684184 Free PMC article.
-
Trichome Independent Resistance against Western Flower Thrips in Tomato.Plant Cell Physiol. 2019 May 1;60(5):1011-1024. doi: 10.1093/pcp/pcz018. Plant Cell Physiol. 2019. PMID: 30715458 Free PMC article.
-
Tomato defences modulate not only insect performance but also their gut microbial composition.Sci Rep. 2023 Oct 24;13(1):18139. doi: 10.1038/s41598-023-44938-2. Sci Rep. 2023. PMID: 37875520 Free PMC article.
-
Arabidopsis CAPRICE (MYB) and GLABRA3 (bHLH) control tomato (Solanum lycopersicum) anthocyanin biosynthesis.PLoS One. 2014 Sep 30;9(9):e109093. doi: 10.1371/journal.pone.0109093. eCollection 2014. PLoS One. 2014. PMID: 25268379 Free PMC article.
-
Morphological characterization of intraspecific variation for trichome traits in tomato (Solanum lycopersicum).Bot Stud. 2023 Mar 29;64(1):7. doi: 10.1186/s40529-023-00370-3. Bot Stud. 2023. PMID: 36988701 Free PMC article.
References
-
- Antonious GF. (2001) Production and quantification of methyl ketones in wild tomato accessions. J Environ Sci Health Part B Pestic Contam Agric Wastes 36: 835–848 - PubMed
-
- Antonious GF, Kochhar TS. (2003) Zingiberene and curcumene in wild tomato. J Environ Sci Health Part B Pestic Contam Agric Wastes 38: 489–500 - PubMed
-
- Antonious GF, Snyder JC. (2006) Natural products: repellency and toxicity of wild tomato leaf extracts to the two-spotted spider mite, Tetranychus urticae Koch. J Environ Sci Health Part B Pestic Contam Agric Wastes 41: 43–55 - PubMed
-
- Aziz N, Paiva NL, May GD, Dixon RA. (2005) Transcriptome analysis of alfalfa glandular trichomes. Planta 221: 28–38 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
