

Proteolytic products of the porcine reproductive and respiratory syndrome virus nsp2 replicase protein
- PMID: 20668084
- PMCID: PMC2937792
- DOI: 10.1128/JVI.01208-10
Proteolytic products of the porcine reproductive and respiratory syndrome virus nsp2 replicase protein
Abstract
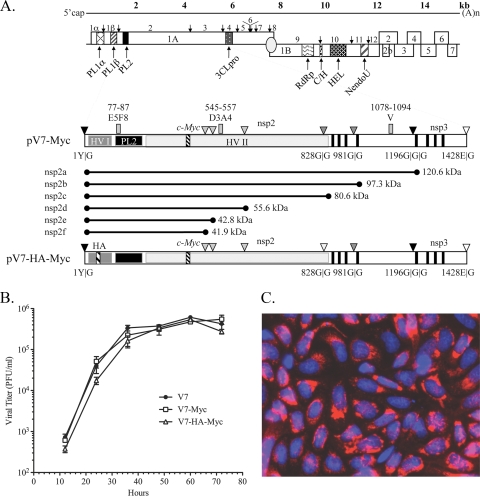
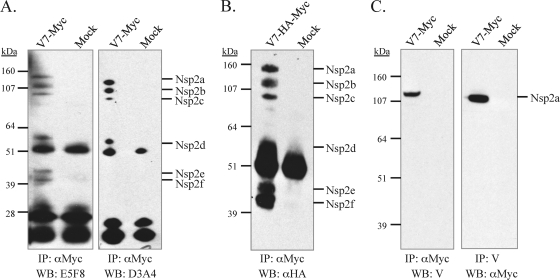
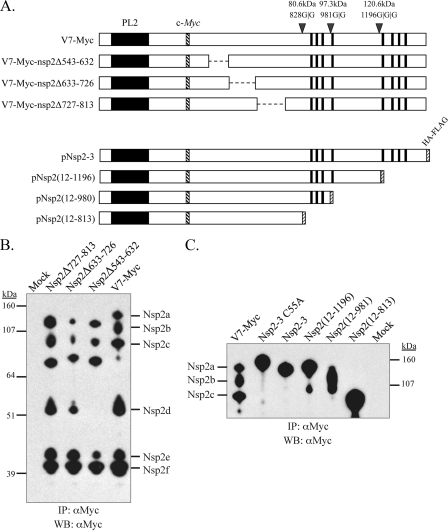
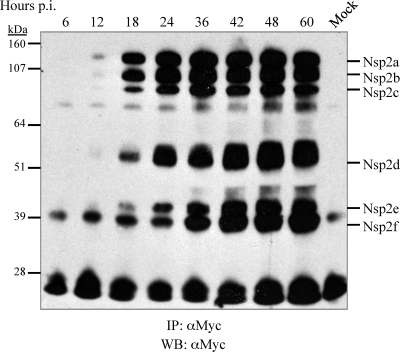
The nsp2 replicase protein of porcine reproductive and respiratory syndrome virus (PRRSV) was recently demonstrated to be processed from its precursor by the PL2 protease at or near the G(1196)|G(1197) dipeptide in transfected CHO cells. Here the proteolytic cleavage of PRRSV nsp2 was further investigated in virally infected MARC-145 cells by using two recombinant PRRSVs expressing epitope-tagged nsp2. The data revealed that PRRSV nsp2 exists as different isoforms, termed nsp2a, nsp2b, nsp2c, nsp2d, nsp2e, and nsp2f, during PRRSV infection. Moreover, on the basis of deletion mutagenesis and antibody probing, these nsp2 species appeared to share the same N terminus but to differ in their C termini. The largest protein, nsp2a, corresponded to the nsp2 product identified in transfected CHO cells. nsp2b and nsp2c were processed within or near the transmembrane (TM) region, presumably at or near the conserved sites G(981)|G(982) and G(828)|G(829)|G(830), respectively. The C termini for nsp2d, -e, and -f were mapped within the nsp2 middle hypervariable region, but no conserved cleavage sites could be definitively predicted. The larger nsp2 species emerged almost simultaneously in the early stage of PRRSV infection. Pulse-chase analysis revealed that all six nsp2 species were relatively stable and had low turnover rates. Deletion mutagenesis revealed that the smaller nsp2 species (e.g., nsp2d, nsp2e, and nsp2f) were not essential for viral replication in cell culture. Lastly, we identified a cellular chaperone, named heat shock 70-kDa protein 5 (HSPA5), that was strongly associated with nsp2, which may have important implications for PRRSV replication. Overall, these findings indicate that PRRSV nsp2 is increasingly emerging as a multifunctional protein and may have a profound impact on PRRSV replication and viral pathogenesis.
Figures


References
-
- Castón, J. R., J. L. Martinez-Torrecuadrada, A. Maraver, E. Lombardo, J. F. Rodriguez, J. I. Casal, and J. L. Carrascosa. 2001. C terminus of infectious bursal disease virus major capsid protein VP2 is involved in definition of the T number for capsid assembly. J. Virol. 75:10815-10828. - PMC - PubMed
-
- Cavanagh, D. 1997. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 142:629-633. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
