

Cellular microRNAs 200b and 429 regulate the Epstein-Barr virus switch between latency and lytic replication
- PMID: 20668090
- PMCID: PMC2937814
- DOI: 10.1128/JVI.00923-10
Cellular microRNAs 200b and 429 regulate the Epstein-Barr virus switch between latency and lytic replication
Abstract
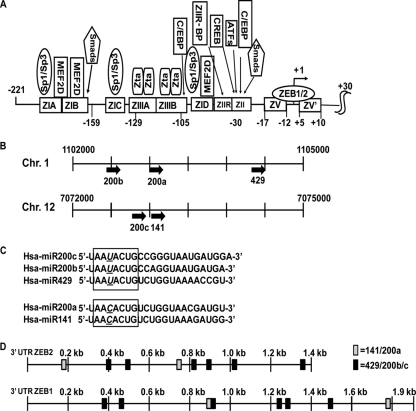
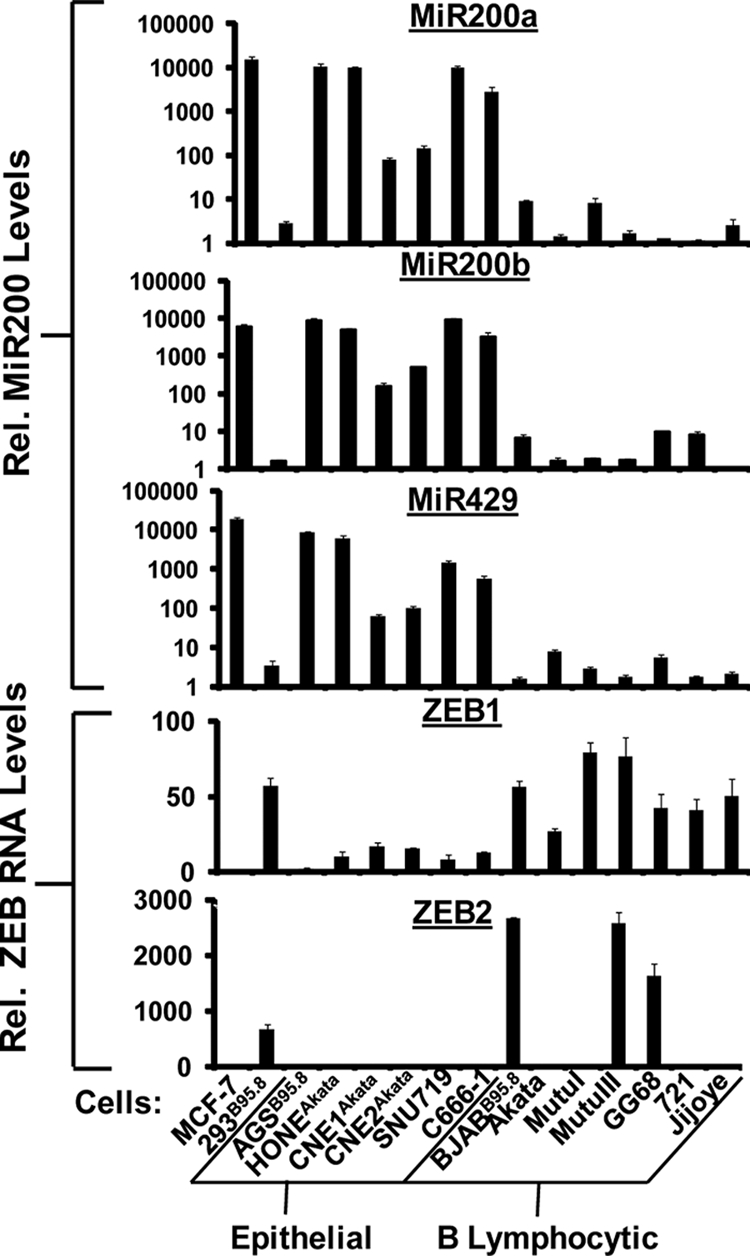
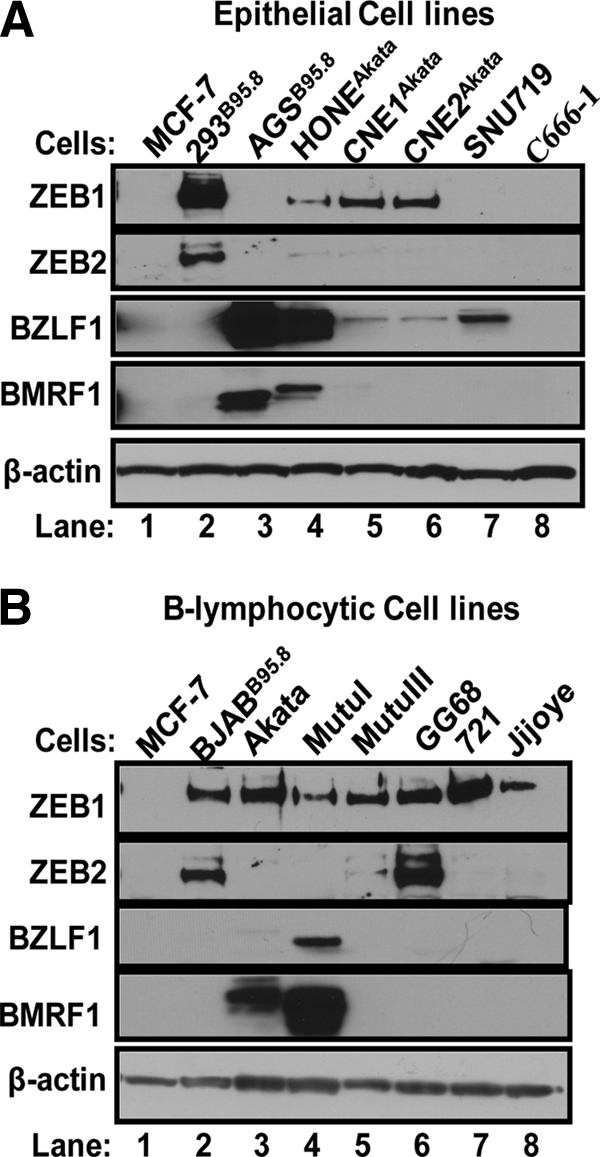
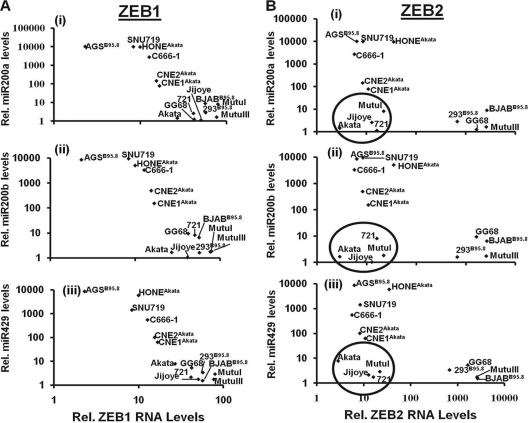
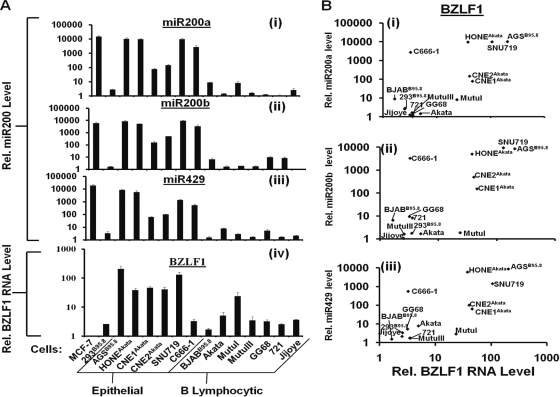
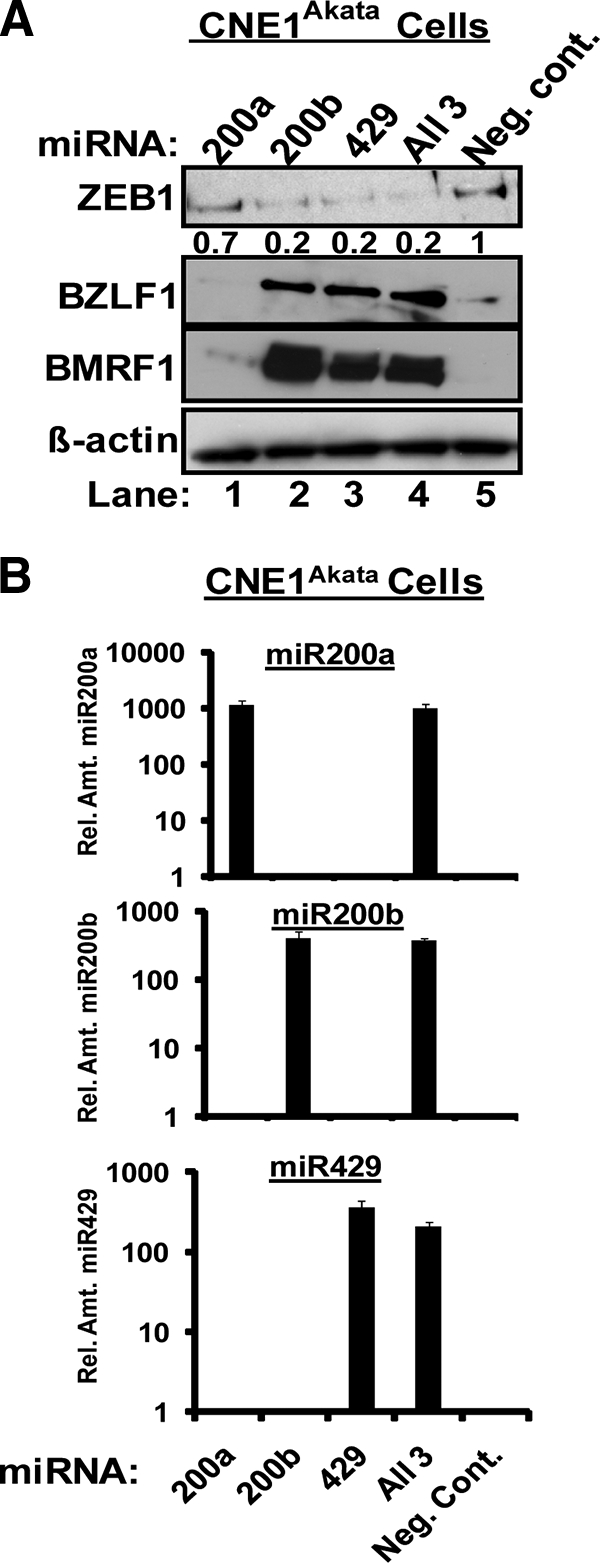
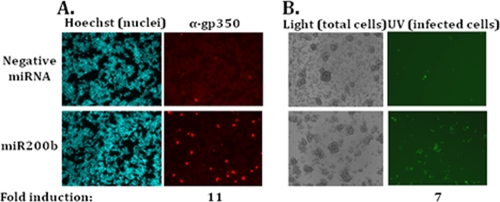
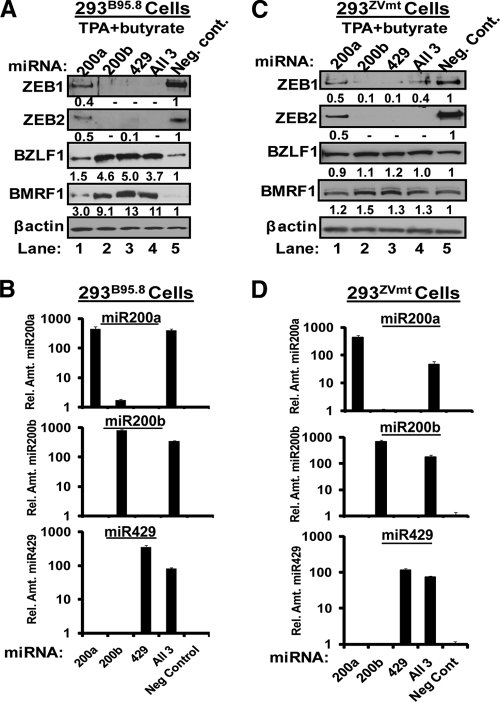
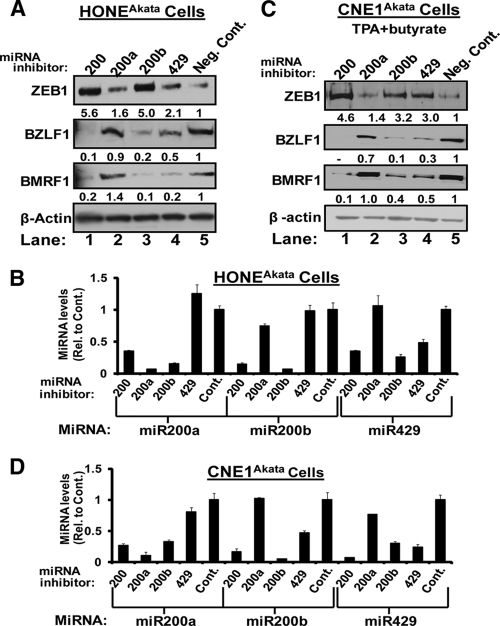
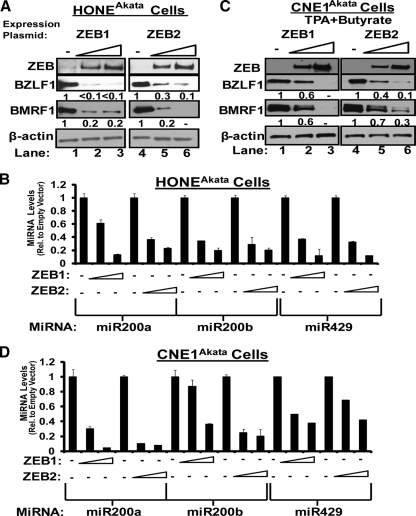
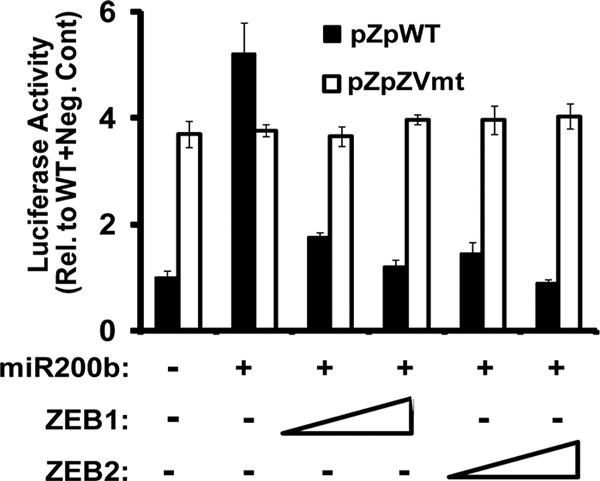
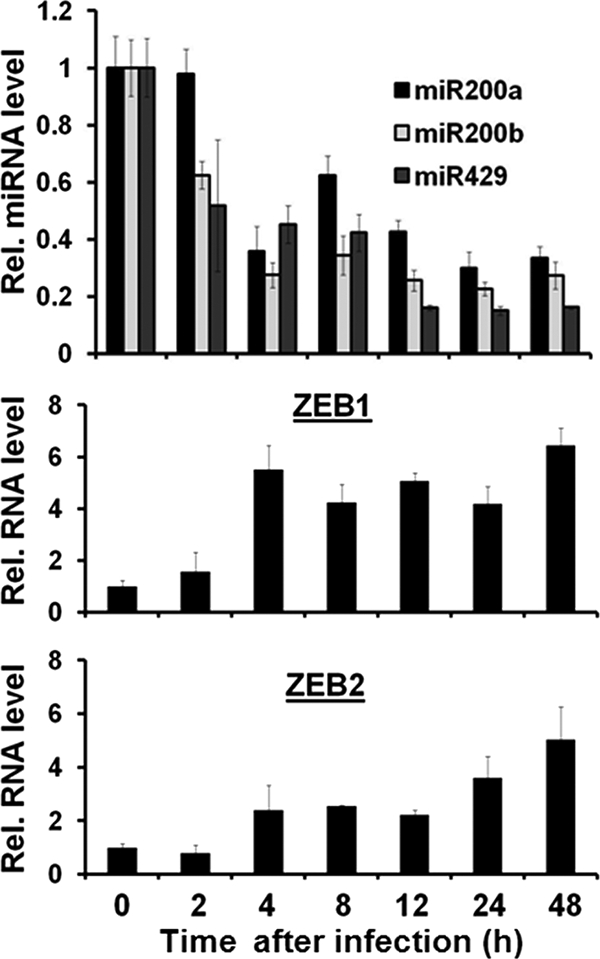
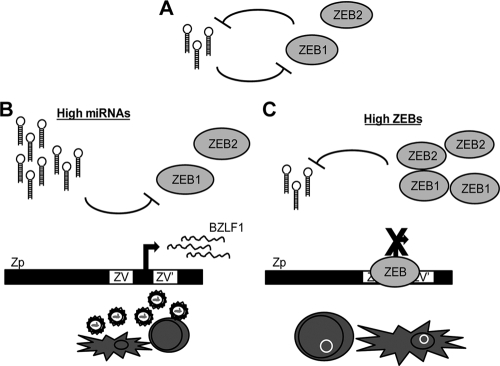
We previously showed that the cellular proteins ZEB1 and ZEB2/SIP1 both play key roles in regulating the latent-lytic switch of Epstein-Barr Virus (EBV) by repressing BZLF1 gene expression. We investigated here the effects of cellular microRNA (miRNA) 200 (miR200) family members on the EBV infection status of cells. We show that miR200b and miR429, but not miR200a, can induce EBV-positive cells into lytic replication by downregulating expression of ZEB1 and ZEB2, leading to production of infectious virus. The levels of miR200 family members in EBV-infected cells strongly negatively correlated with the levels of the ZEBs (e.g., -0.89 [P < 0.001] for miR429 versus ZEB1) and positively correlated with the degree of EBV lytic gene expression (e.g., 0.73 [P < 0.01] for miR429 versus BZLF1). The addition of either miR200b or miR429 to EBV-positive cells led to EBV lytic reactivation in a ZEB-dependent manner; inhibition of these miRNAs led to decreased EBV lytic gene expression. The degree of latent infection by an EBV mutant defective in the primary ZEB-binding site of the EBV BZLF1 promoter was not affected by the addition of these miRNAs. Furthermore, EBV infection of primary blood B cells led to downregulation of these miRNAs and upregulation of ZEB levels. Thus, we conclude that miRNAs 200b and 429 are key regulators via their effects on expression of ZEB1 and ZEB2 of the switch between latent and lytic infection by EBV and, therefore, potential targets for development of new lytic induction therapeutics with which to treat patients with EBV-associated malignancies.
Figures
References
-
- Adam, L., M. Zhong, W. Choi, W. Qi, M. Nicoloso, A. Arora, G. Calin, H. Wang, A. Siefker-Radtke, D. McConkey, M. Bar-Eli, and C. Dinney. 2009. miR-200 expression regulates epithelial-to-mesenchymal transition in bladder cancer cells and reverses resistance to epidermal growth factor receptor therapy. Clin. Cancer Res. 15:5060-5072. - PMC - PubMed
-
- Ambros, V. 2000. Control of developmental timing in Caenorhabditis elegans. Curr. Opin. Genet. Dev. 10:428-433. - PubMed
-
- Bartel, D. P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116:281-297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
