

Extracellular ATP may contribute to tissue repair by rapidly stimulating purinergic receptor X7-dependent vascular endothelial growth factor release from primary human monocytes
- PMID: 20668222
- PMCID: PMC3156583
- DOI: 10.4049/jimmunol.1001298
Extracellular ATP may contribute to tissue repair by rapidly stimulating purinergic receptor X7-dependent vascular endothelial growth factor release from primary human monocytes
Abstract
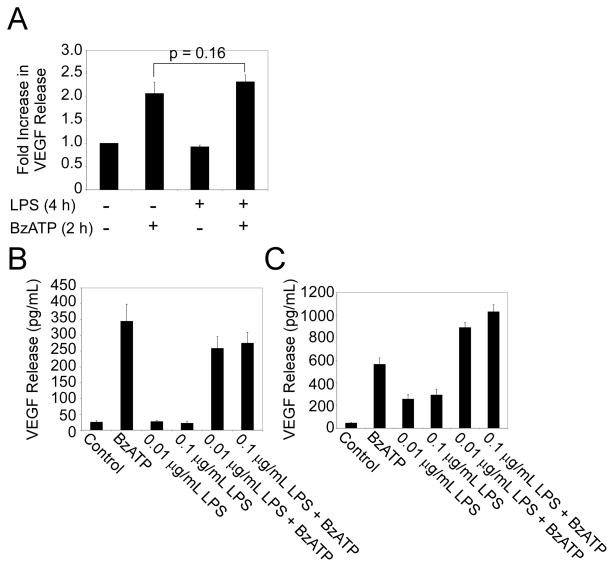
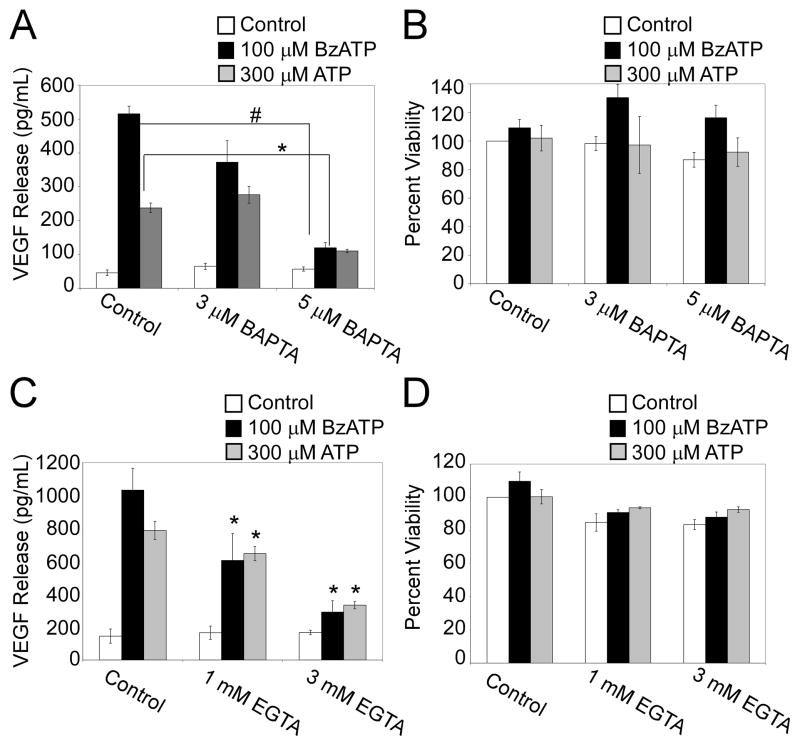
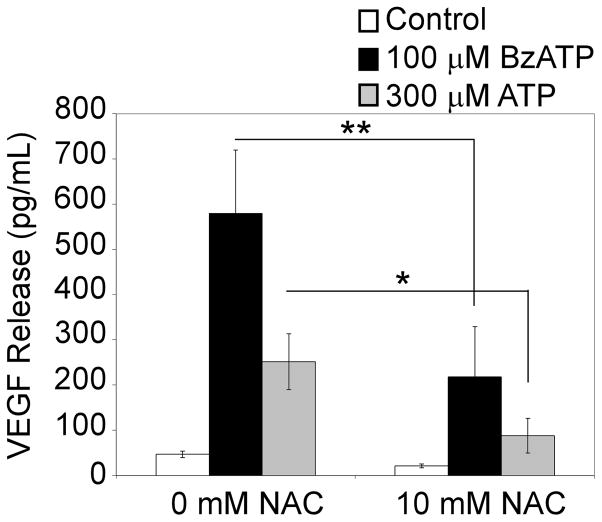
Extracellular ATP has been proposed to act as a danger signal to alert the immune system of cell damage. Release of high local concentrations of ATP activates the nucleotide receptor, purinergic receptor X7 (P2RX7), on monocytic cells, which promotes the processing/release of proinflammatory mediators. Although the proinflammatory actions of P2RX7 are well recognized, little is known regarding the potential function of P2RX7 in repair responses. Because the resolution of inflammation is characterized by monocytic cell-dependent production of proangiogenic factors, we evaluated the contribution of P2RX7 to this process. We observed that both short-term and long-term P2RX7 activation promotes the robust release of vascular endothelial growth factor from primary human monocytes. This vascular endothelial growth factor release is calcium dependent and associated with reactive oxygen species production. This previously unrecognized action of P2RX7 suggests that it may not only participate in inflammation and cell death, but that it is also likely to be important in the control of angiogenesis and wound repair.
Figures
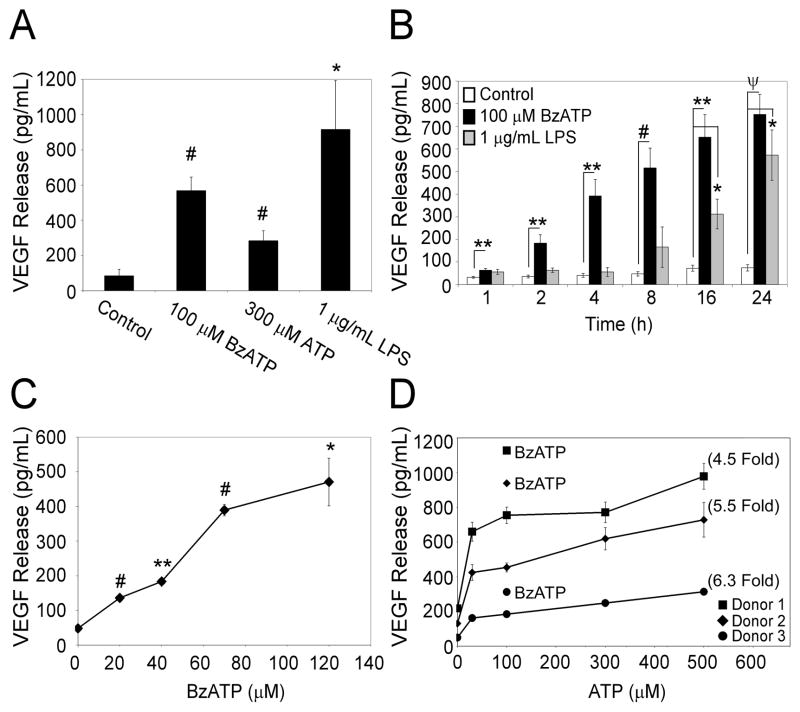
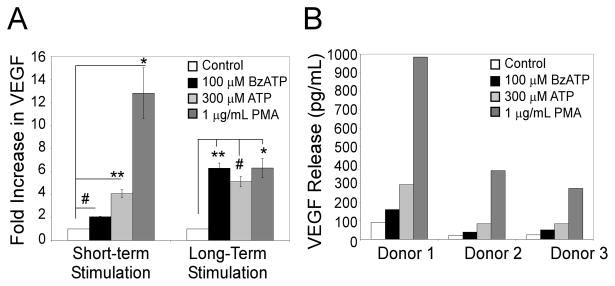
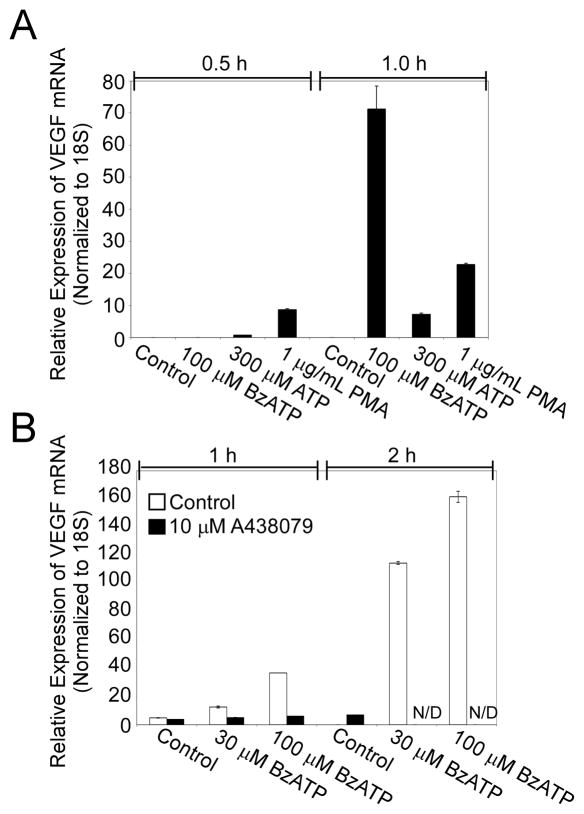
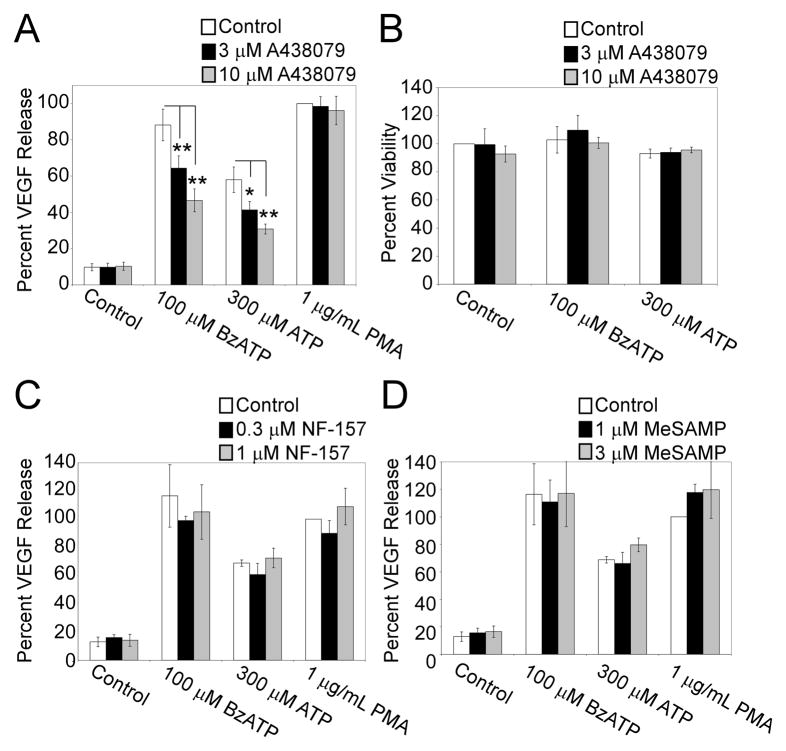
Similar articles
-
Activation of the transcription factor FosB/activating protein-1 (AP-1) is a prominent downstream signal of the extracellular nucleotide receptor P2RX7 in monocytic and osteoblastic cells.J Biol Chem. 2010 Oct 29;285(44):34288-98. doi: 10.1074/jbc.M110.142091. Epub 2010 Sep 2. J Biol Chem. 2010. PMID: 20813842 Free PMC article.
-
Extracellular ATP acting at the P2X7 receptor inhibits secretion of soluble HLA-G from human monocytes.J Immunol. 2009 Oct 1;183(7):4302-11. doi: 10.4049/jimmunol.0804265. Epub 2009 Sep 11. J Immunol. 2009. PMID: 19748989
-
Purinergic receptor modulation of lipopolysaccharide signaling and inducible nitric-oxide synthase expression in RAW 264.7 macrophages.J Biol Chem. 1998 Oct 16;273(42):27170-5. doi: 10.1074/jbc.273.42.27170. J Biol Chem. 1998. PMID: 9765236
-
Purinergic signaling in kidney disease.Kidney Int. 2017 Feb;91(2):315-323. doi: 10.1016/j.kint.2016.08.029. Epub 2016 Oct 22. Kidney Int. 2017. PMID: 27780585 Review.
-
A new approach to the inflammatory/autoimmune diseases.Recent Pat Antiinfect Drug Discov. 2009 Jun;4(2):108-13. doi: 10.2174/157489109788490343. Recent Pat Antiinfect Drug Discov. 2009. PMID: 19519545 Review.
Cited by
-
Activation of P2X7 and P2Y11 purinergic receptors inhibits migration and normalizes tumor-derived endothelial cells via cAMP signaling.Sci Rep. 2016 Sep 2;6:32602. doi: 10.1038/srep32602. Sci Rep. 2016. PMID: 27586846 Free PMC article.
-
The Role of Autophagy in the Resistance to BRAF Inhibition in BRAF-Mutated Melanoma.Target Oncol. 2018 Aug;13(4):437-446. doi: 10.1007/s11523-018-0565-2. Target Oncol. 2018. PMID: 29667105 Review.
-
Emerging Role for Ferroptosis in Infectious Diseases.Adv Exp Med Biol. 2021;1301:59-79. doi: 10.1007/978-3-030-62026-4_5. Adv Exp Med Biol. 2021. PMID: 34370288 Review.
-
An autophagy-driven pathway of ATP secretion supports the aggressive phenotype of BRAFV600E inhibitor-resistant metastatic melanoma cells.Autophagy. 2017 Sep 2;13(9):1512-1527. doi: 10.1080/15548627.2017.1332550. Epub 2017 Jul 19. Autophagy. 2017. PMID: 28722539 Free PMC article.
-
Lack of functional P2X7 receptor aggravates brain edema development after middle cerebral artery occlusion.Purinergic Signal. 2016 Sep;12(3):453-63. doi: 10.1007/s11302-016-9511-x. Epub 2016 Apr 5. Purinergic Signal. 2016. PMID: 27048203 Free PMC article.
References
-
- Rubartelli A, Lotze MT. Inside, outside, upside down: damage-associated molecular-pattern molecules (DAMPs) and redox. Trends Immunol. 2007;28:429–436. - PubMed
-
- la Sala A, Ferrari D, Di Virgilio F, Idzko M, Norgauer J, Girolomoni G. Alerting and tuning the immune response by extracellular nucleotides. J Leukoc Biol. 2003;73:339–343. - PubMed
-
- Labasi JM, Petrushova N, Donovan C, McCurdy S, Lira P, Payette MM, Brissette W, Wicks JR, Audoly L, Gabel CA. Absence of the P2X7 receptor alters leukocyte function and attenuates an inflammatory response. J Immunol. 2002;168:6436–6445. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
