

Modes and models of forebrain cholinergic neuromodulation of cognition
- PMID: 20668433
- PMCID: PMC2992803
- DOI: 10.1038/npp.2010.104
Modes and models of forebrain cholinergic neuromodulation of cognition
Abstract
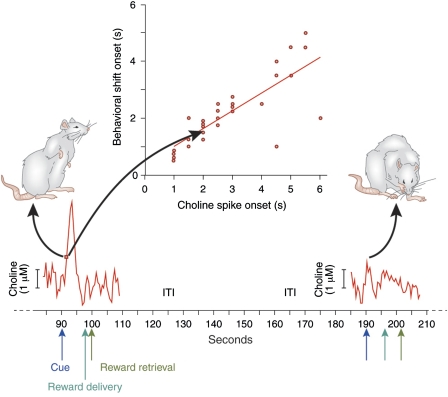
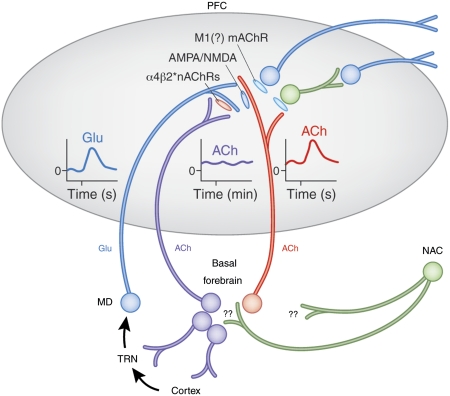
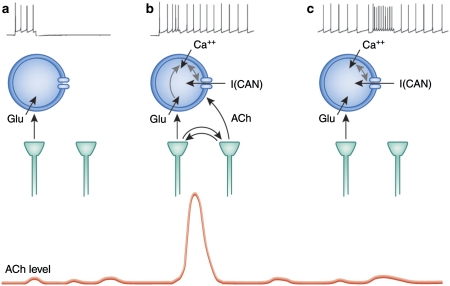
As indicated by the profound cognitive impairments caused by cholinergic receptor antagonists, cholinergic neurotransmission has a vital role in cognitive function, specifically attention and memory encoding. Abnormally regulated cholinergic neurotransmission has been hypothesized to contribute to the cognitive symptoms of neuropsychiatric disorders. Loss of cholinergic neurons enhances the severity of the symptoms of dementia. Cholinergic receptor agonists and acetylcholinesterase inhibitors have been investigated for the treatment of cognitive dysfunction. Evidence from experiments using new techniques for measuring rapid changes in cholinergic neurotransmission provides a novel perspective on the cholinergic regulation of cognitive processes. This evidence indicates that changes in cholinergic modulation on a timescale of seconds is triggered by sensory input cues and serves to facilitate cue detection and attentional performance. Furthermore, the evidence indicates cholinergic induction of evoked intrinsic, persistent spiking mechanisms for active maintenance of sensory input, and planned responses. Models have been developed to describe the neuronal mechanisms underlying the transient modulation of cortical target circuits by cholinergic activity. These models postulate specific locations and roles of nicotinic and muscarinic acetylcholine receptors and that cholinergic neurotransmission is controlled in part by (cortical) target circuits. The available evidence and these models point to new principles governing the development of the next generation of cholinergic treatments for cognitive disorders.
Figures
References
-
- Aarsland D, Larsen JP, Reinvang I, Aasland AM. Effects of cholinergic blockade on language in healthy young women. Brain. 1994;117:1377–1384. - PubMed
-
- Aggleton JP, Brown MW. Episodic memory, amnesia, and the hippocampal-anterior thalamic axis. Behav Brain Sci. 1999;22:425–444. - PubMed
-
- Aigner TG, Mishkin M. The effects of physostigmine and scopolamine on recognition memory in monkeys. Behav Neurosci. 1986;45:81–87. - PubMed
-
- Aigner TG, Mitchell SJ, Aggleton JP, DeLong MR, Struble RG, Price DL, et al. Effects of scopolamine and physostigmine on recognition memory in monkeys with ibotenic-acid lesions of the nucleus basalis of Meynert. Psychopharmacology. 1987;92:292–300. - PubMed
-
- Aigner TG, Walker DL, Mishkin M. Comparison of the effects of scopolamine administered before and after acquisition in a test of visual recognition memory in monkeys. Behav Neural Biol. 1991;55:61–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
