

NRMT is an alpha-N-methyltransferase that methylates RCC1 and retinoblastoma protein
- PMID: 20668449
- PMCID: PMC2939154
- DOI: 10.1038/nature09343
NRMT is an alpha-N-methyltransferase that methylates RCC1 and retinoblastoma protein
Abstract
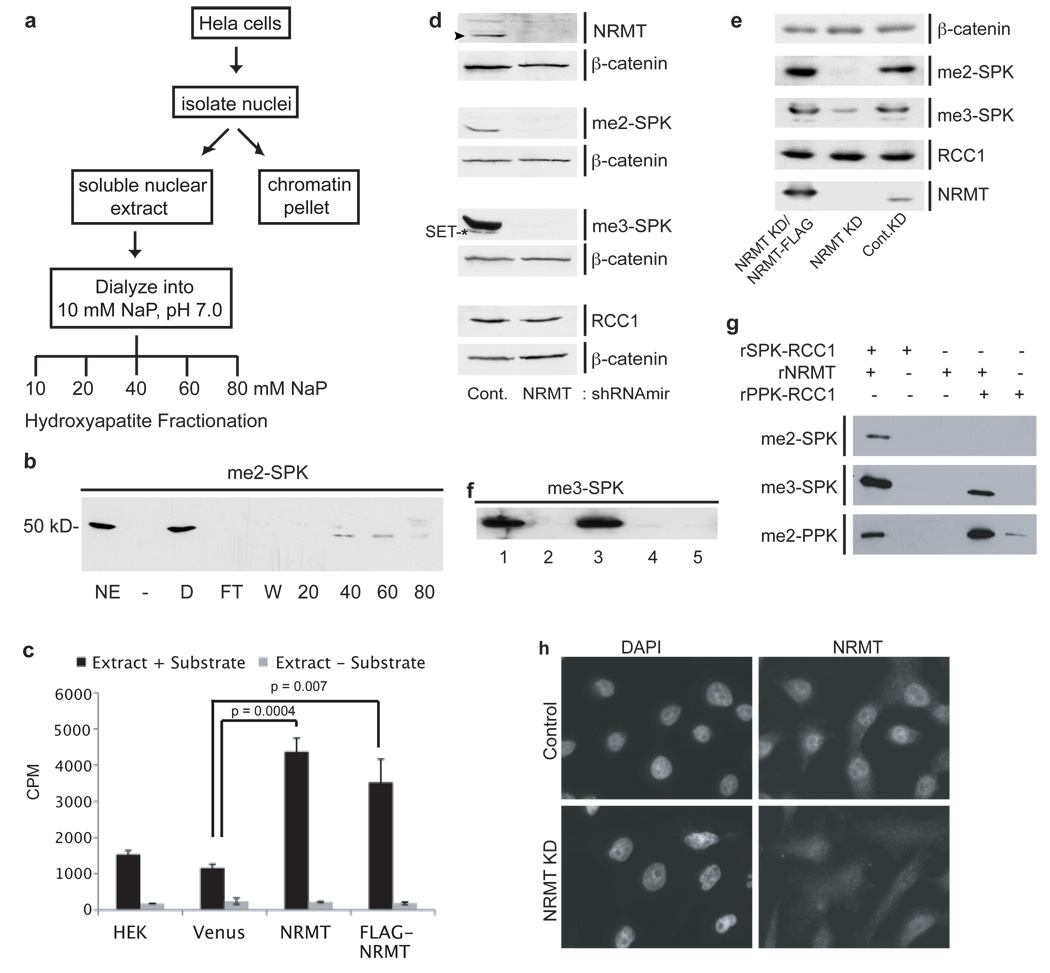
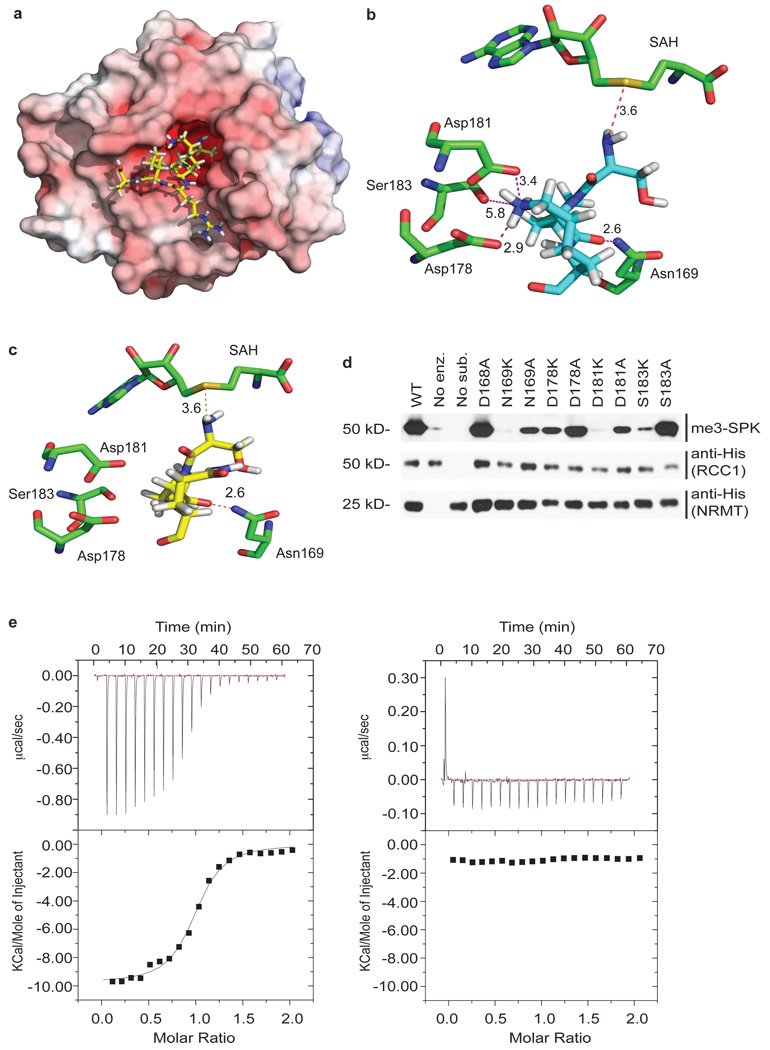
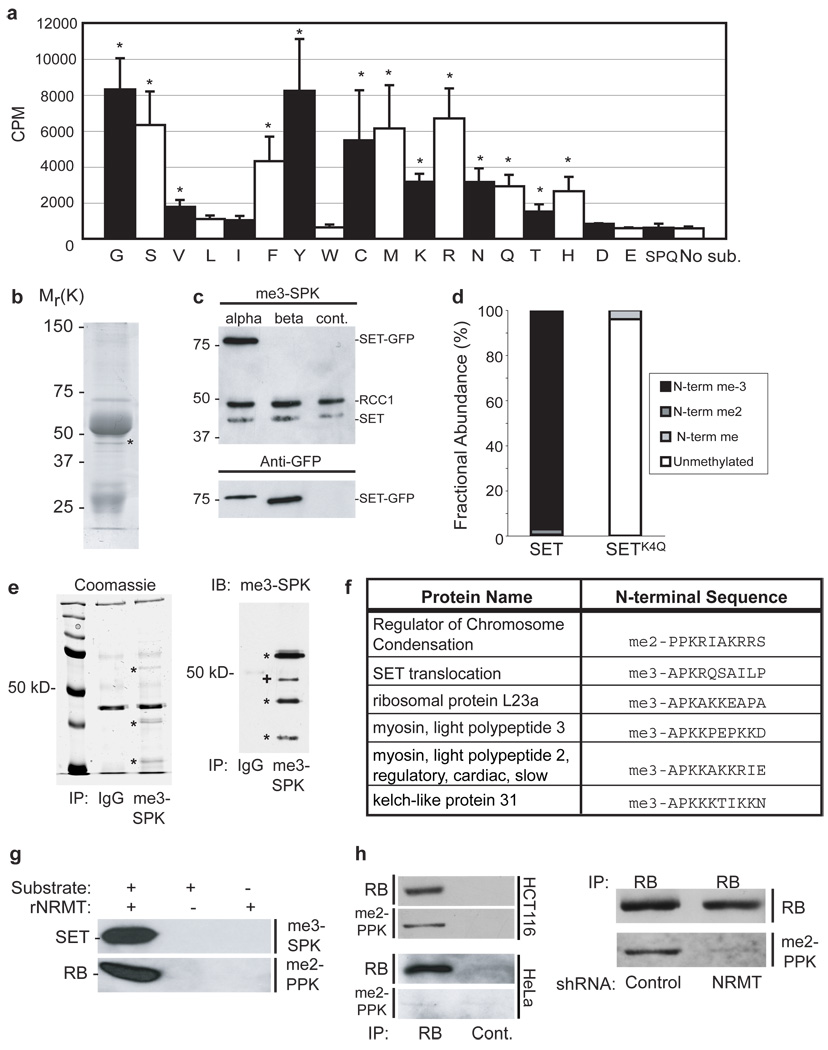
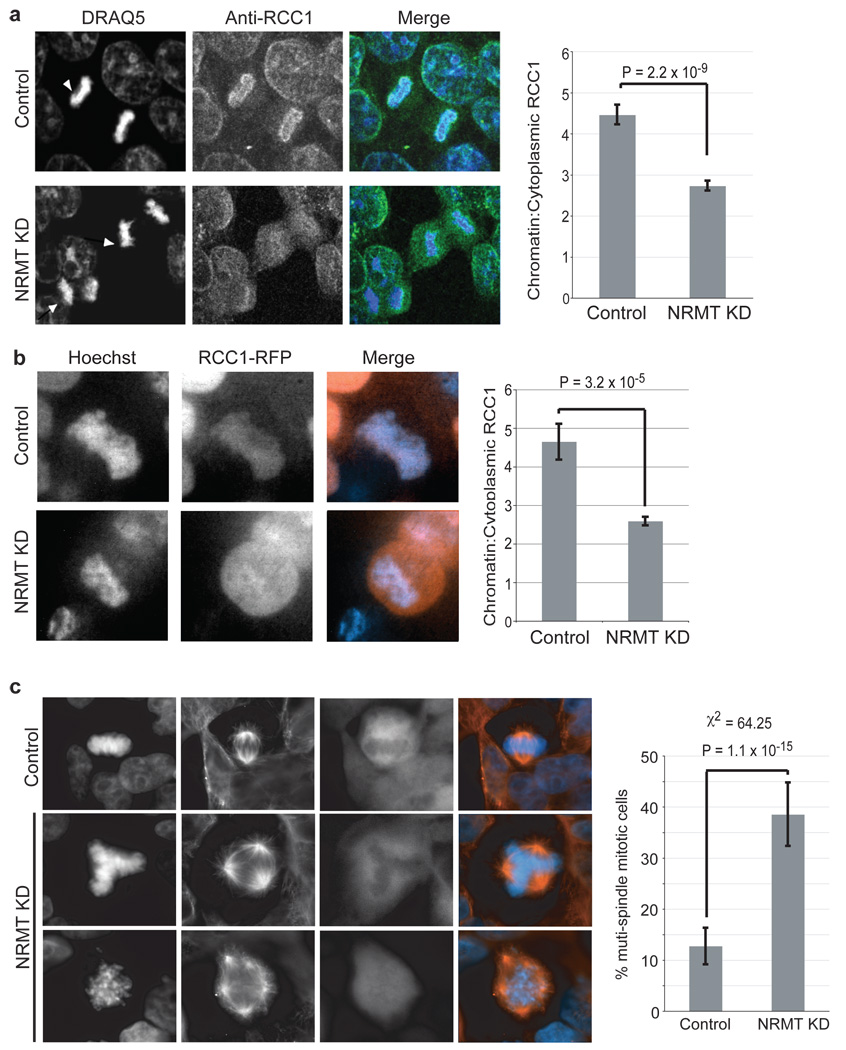
The post-translational methylation of alpha-amino groups was first discovered over 30 years ago on the bacterial ribosomal proteins L16 and L33 (refs 1, 2), but almost nothing is known about the function or enzymology of this modification. Several other bacterial and eukaryotic proteins have since been shown to be alpha-N-methylated. However, the Ran guanine nucleotide-exchange factor, RCC1, is the only protein for which any biological function of alpha-N-methylation has been identified. Methylation-defective mutants of RCC1 have reduced affinity for DNA and cause mitotic defects, but further characterization of this modification has been hindered by ignorance of the responsible methyltransferase. All fungal and animal N-terminally methylated proteins contain a unique N-terminal motif, Met-(Ala/Pro/Ser)-Pro-Lys, indicating that they may be targets of the same, unknown enzyme. The initiating Met is cleaved, and the exposed alpha-amino group is mono-, di- or trimethylated. Here we report the discovery of the first alpha-N-methyltransferase, which we named N-terminal RCC1 methyltransferase (NRMT). Substrate docking and mutational analysis of RCC1 defined the NRMT recognition sequence and enabled the identification of numerous new methylation targets, including SET (also known as TAF-I or PHAPII) and the retinoblastoma protein, RB. Knockdown of NRMT recapitulates the multi-spindle phenotype seen with methylation-defective RCC1 mutants, demonstrating the importance of alpha-N-methylation for normal bipolar spindle formation and chromosome segregation.
Figures
Similar articles
-
The methylated N-terminal tail of RCC1 is required for stabilisation of its interaction with chromatin by Ran in live cells.BMC Cell Biol. 2010 Jun 21;11:43. doi: 10.1186/1471-2121-11-43. BMC Cell Biol. 2010. PMID: 20565941 Free PMC article.
-
N-terminal alpha-methylation of RCC1 is necessary for stable chromatin association and normal mitosis.Nat Cell Biol. 2007 May;9(5):596-603. doi: 10.1038/ncb1572. Epub 2007 Apr 15. Nat Cell Biol. 2007. PMID: 17435751 Free PMC article.
-
Substrate specificity of mammalian N-terminal α-amino methyltransferase NRMT.Biochemistry. 2012 Jul 31;51(30):5942-50. doi: 10.1021/bi300278f. Epub 2012 Jul 19. Biochemistry. 2012. PMID: 22769851 Free PMC article.
-
The intricate roles of RCC1 in normal cells and cancer cells.Biochem Soc Trans. 2022 Feb 28;50(1):83-93. doi: 10.1042/BST20210861. Biochem Soc Trans. 2022. PMID: 35191966 Review.
-
RCC1 in the Ran pathway.J Biochem. 1996 Aug;120(2):207-14. doi: 10.1093/oxfordjournals.jbchem.a021400. J Biochem. 1996. PMID: 8889801 Review.
Cited by
-
N-terminal acetylation and methylation differentially affect the function of MYL9.Biochem J. 2018 Oct 22;475(20):3201-3219. doi: 10.1042/BCJ20180638. Biochem J. 2018. PMID: 30242065 Free PMC article.
-
The ribosomal l1 protuberance in yeast is methylated on a lysine residue catalyzed by a seven-beta-strand methyltransferase.J Biol Chem. 2011 May 27;286(21):18405-13. doi: 10.1074/jbc.M110.200410. Epub 2011 Apr 1. J Biol Chem. 2011. PMID: 21460220 Free PMC article.
-
Optimizing purification and activity assays of N-terminal methyltransferase complexes.Methods Enzymol. 2023;684:71-111. doi: 10.1016/bs.mie.2023.02.001. Epub 2023 Mar 6. Methods Enzymol. 2023. PMID: 37230594 Free PMC article.
-
Posttranslational modification of CENP-A influences the conformation of centromeric chromatin.Proc Natl Acad Sci U S A. 2013 Jul 16;110(29):11827-32. doi: 10.1073/pnas.1300325110. Epub 2013 Jul 1. Proc Natl Acad Sci U S A. 2013. PMID: 23818633 Free PMC article.
-
Probing the Plasticity in the Active Site of Protein N-terminal Methyltransferase 1 Using Bisubstrate Analogues.J Med Chem. 2020 Aug 13;63(15):8419-8431. doi: 10.1021/acs.jmedchem.0c00770. Epub 2020 Jul 16. J Med Chem. 2020. PMID: 32605369 Free PMC article.
References
-
- Brosius J, Chen R. The primary structure of protein L16 located at the peptidyltransferase center of Escherichia coli ribosomes. FEBS Lett. 1976;68:105–109. - PubMed
-
- Wittmann-Liebold B, Pannenbecker R. Primary structure of protein L33 from the large subunit of the Escherichia coli ribosome. FEBS Lett. 1976;68:115–118. - PubMed
-
- Meng F, et al. Molecular-level description of proteins from saccharomyces cerevisiae using quadrupole FT hybrid mass spectrometry for top down proteomics. Anal Chem. 2004;76:2852–2858. - PubMed
-
- Sadaie M, Shinmyozu K, Nakayama J. A conserved SET domain methyltransferase, Set11, modifies ribosomal protein Rpl12 in fission yeast. J Biol Chem. 2008;283:7185–7195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
