

KRIT1 regulates the homeostasis of intracellular reactive oxygen species
- PMID: 20668652
- PMCID: PMC2910502
- DOI: 10.1371/journal.pone.0011786
KRIT1 regulates the homeostasis of intracellular reactive oxygen species
Erratum in
-
Correction: KRIT1 Regulates the Homeostasis of Intracellular Reactive Oxygen Species.PLoS One. 2019 Nov 7;14(11):e0223089. doi: 10.1371/journal.pone.0223089. eCollection 2019. PLoS One. 2019. PMID: 31697685 Free PMC article.
Abstract
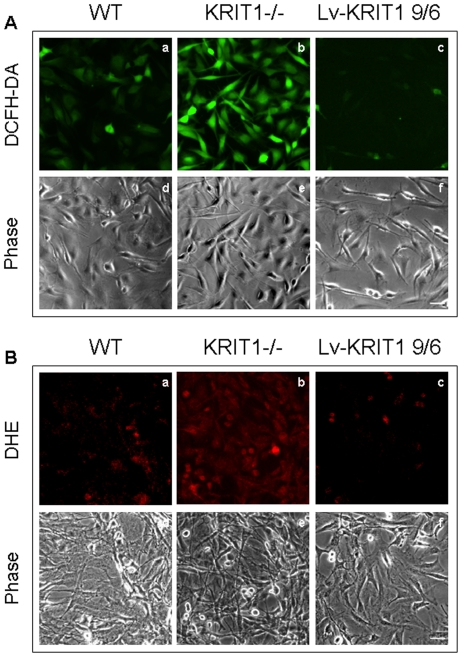
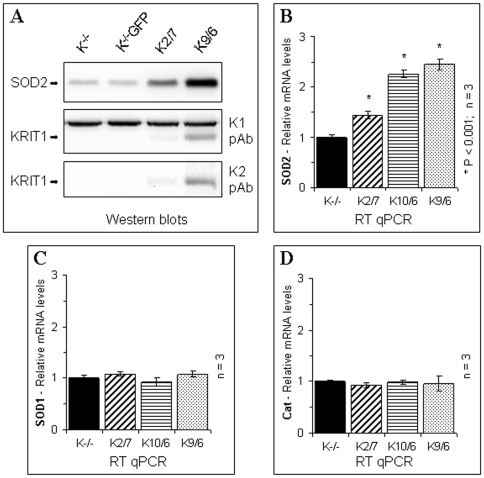
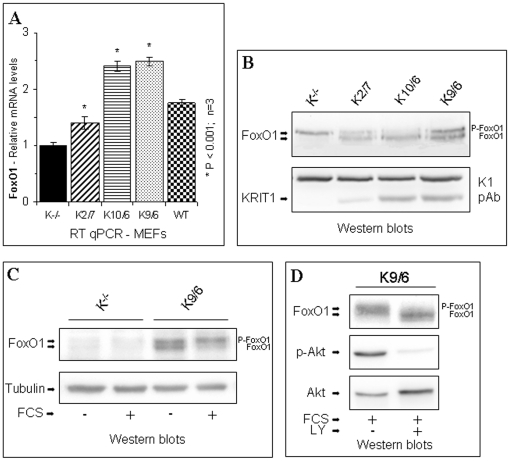
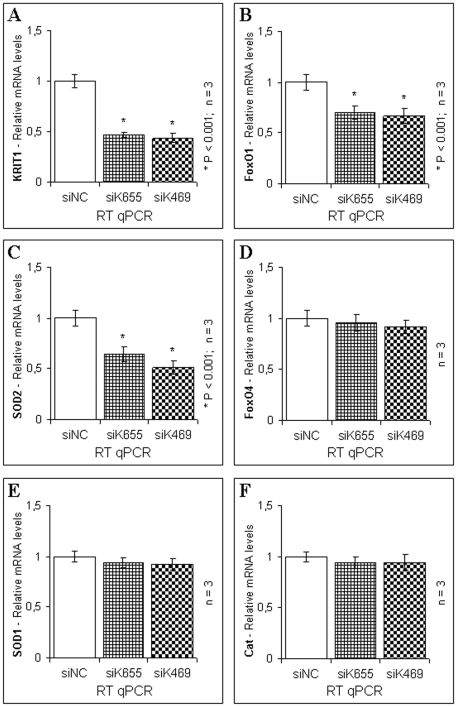
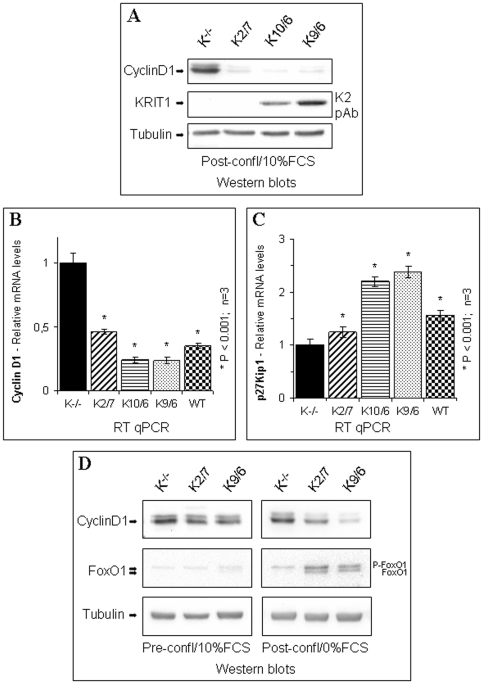
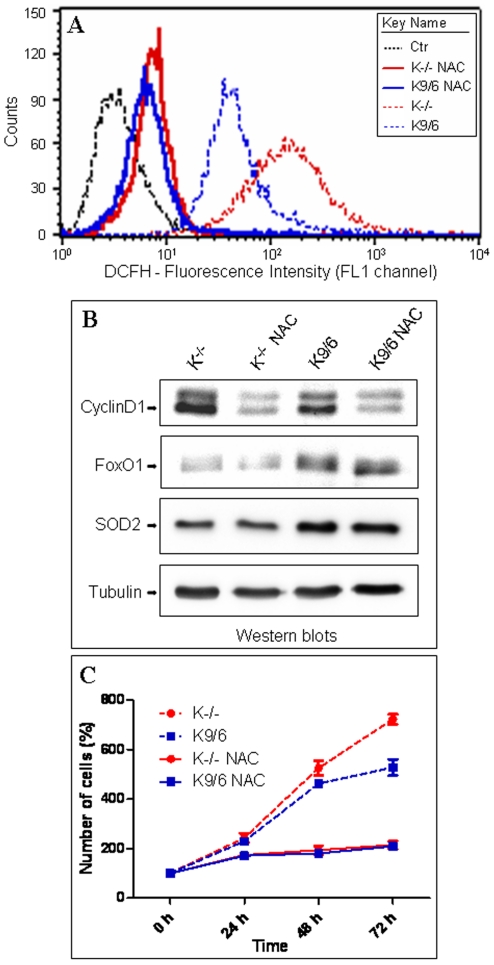
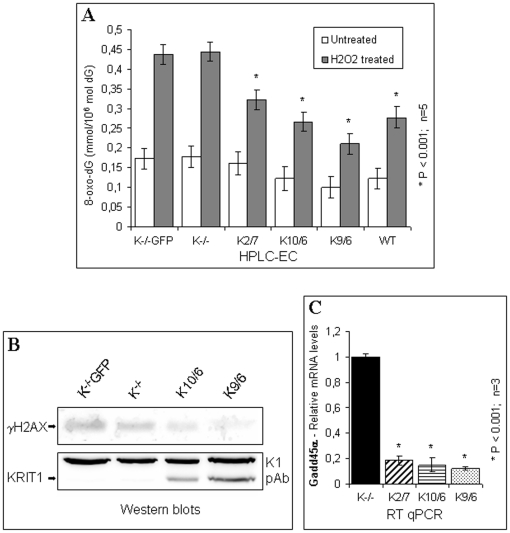
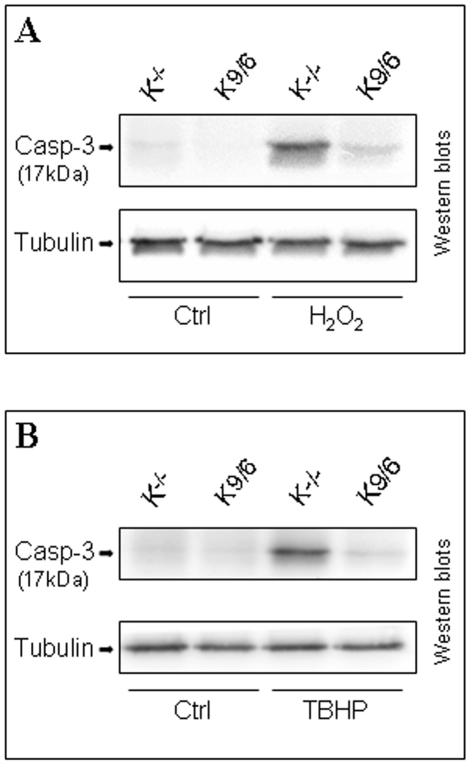
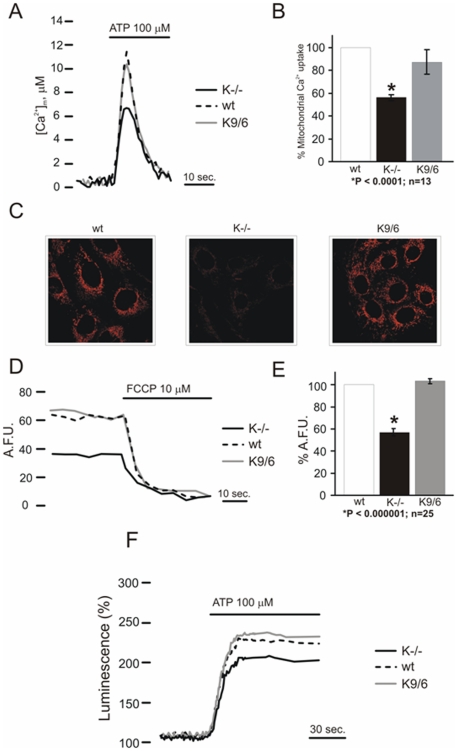
KRIT1 is a gene responsible for Cerebral Cavernous Malformations (CCM), a major cerebrovascular disease characterized by abnormally enlarged and leaky capillaries that predispose to seizures, focal neurological deficits, and fatal intracerebral hemorrhage. Comprehensive analysis of the KRIT1 gene in CCM patients has suggested that KRIT1 functions need to be severely impaired for pathogenesis. However, the molecular and cellular functions of KRIT1 as well as CCM pathogenesis mechanisms are still research challenges. We found that KRIT1 plays an important role in molecular mechanisms involved in the maintenance of the intracellular Reactive Oxygen Species (ROS) homeostasis to prevent oxidative cellular damage. In particular, we demonstrate that KRIT1 loss/down-regulation is associated with a significant increase in intracellular ROS levels. Conversely, ROS levels in KRIT1(-/-) cells are significantly and dose-dependently reduced after restoration of KRIT1 expression. Moreover, we show that the modulation of intracellular ROS levels by KRIT1 loss/restoration is strictly correlated with the modulation of the expression of the antioxidant protein SOD2 as well as of the transcriptional factor FoxO1, a master regulator of cell responses to oxidative stress and a modulator of SOD2 levels. Furthermore, we show that the KRIT1-dependent maintenance of low ROS levels facilitates the downregulation of cyclin D1 expression required for cell transition from proliferative growth to quiescence. Finally, we demonstrate that the enhanced ROS levels in KRIT1(-/-) cells are associated with an increased cell susceptibility to oxidative DNA damage and a marked induction of the DNA damage sensor and repair gene Gadd45alpha, as well as with a decline of mitochondrial energy metabolism. Taken together, our results point to a new model where KRIT1 limits the accumulation of intracellular oxidants and prevents oxidative stress-mediated cellular dysfunction and DNA damage by enhancing the cell capacity to scavenge intracellular ROS through an antioxidant pathway involving FoxO1 and SOD2, thus providing novel and useful insights into the understanding of KRIT1 molecular and cellular functions.
Conflict of interest statement
Figures
References
-
- Serebriiskii I, Estojak J, Sonoda G, Testa JR, Golemis EA. Association of Krev-1/rap1a with Krit1, a novel ankyrin repeat-containing protein encoded by a gene mapping to 7q21-22. Oncogene. 1997;15:1043–1049. - PubMed
-
- Labauge P, Denier C, Bergametti F, Tournier-Lasserve E. Genetics of cavernous angiomas. Lancet Neurol. 2007;6:237–244. - PubMed
-
- Laberge-le Couteulx S, Jung HH, Labauge P, Houtteville JP, Lescoat C, et al. Truncating mutations in CCM1, encoding KRIT1, cause hereditary cavernous angiomas. Nat Genet. 1999;23:189–193. - PubMed
-
- Sahoo T, Johnson EW, Thomas JW, Kuehl PM, Jones TL, et al. Mutations in the gene encoding KRIT1, a Krev-1/rap1a binding protein, cause cerebral cavernous malformations (CCM1). Hum Mol Genet. 1999;8:2325–2333. - PubMed
-
- Marchuk DA, Srinivasan S, Squire TL, Zawistowski JS. Vascular morphogenesis: tales of two syndromes. Hum Mol Genet. 2003;12 Spec No 1:R97–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
