

Structure of the CaMKIIdelta/calmodulin complex reveals the molecular mechanism of CaMKII kinase activation
- PMID: 20668654
- PMCID: PMC2910593
- DOI: 10.1371/journal.pbio.1000426
Structure of the CaMKIIdelta/calmodulin complex reveals the molecular mechanism of CaMKII kinase activation
Abstract
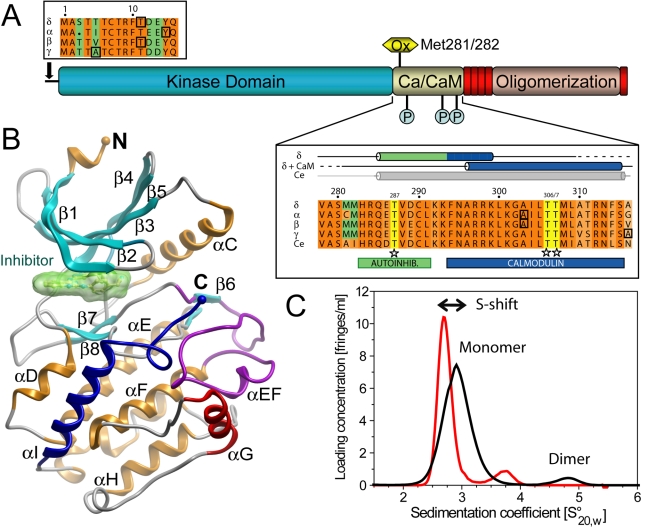
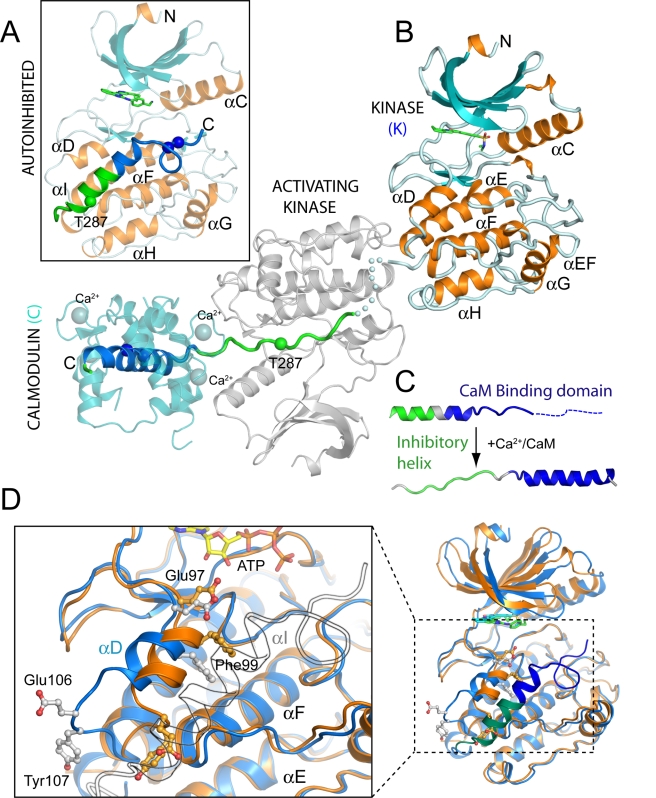
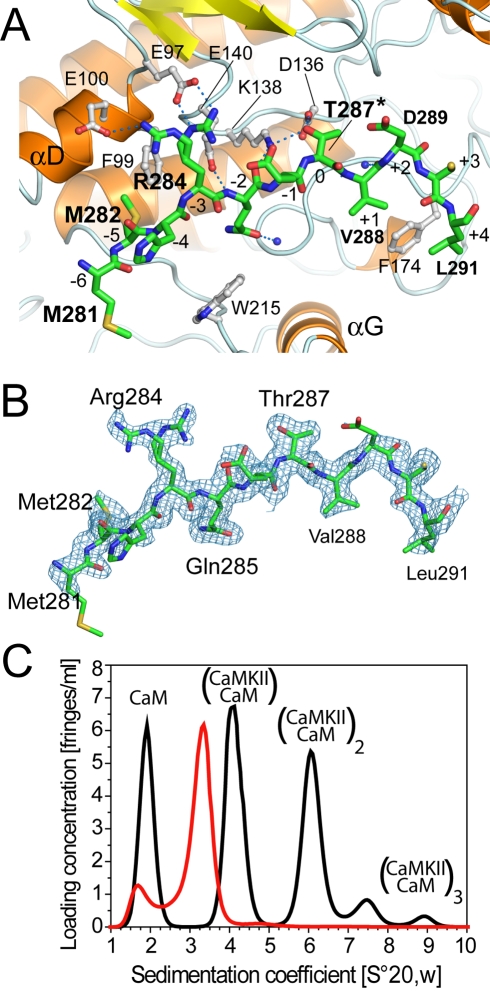
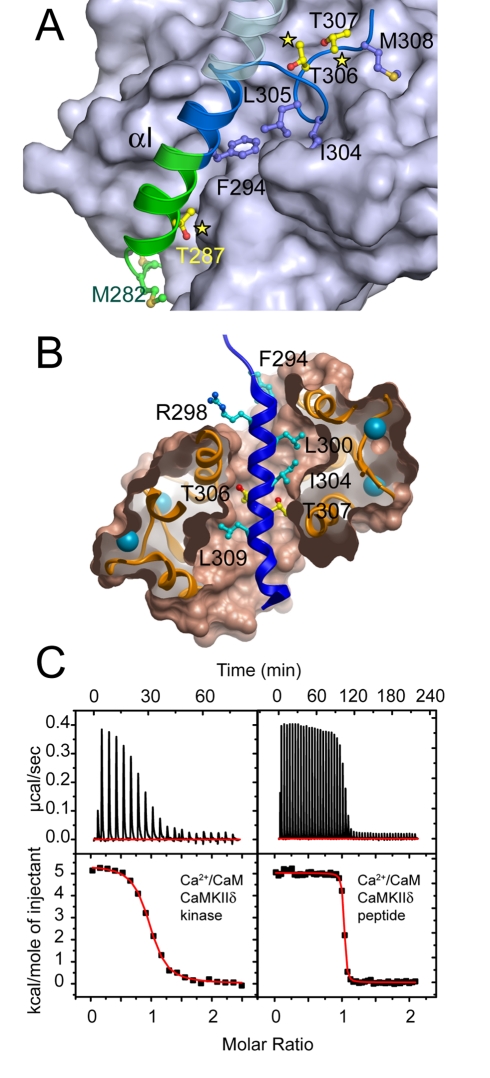
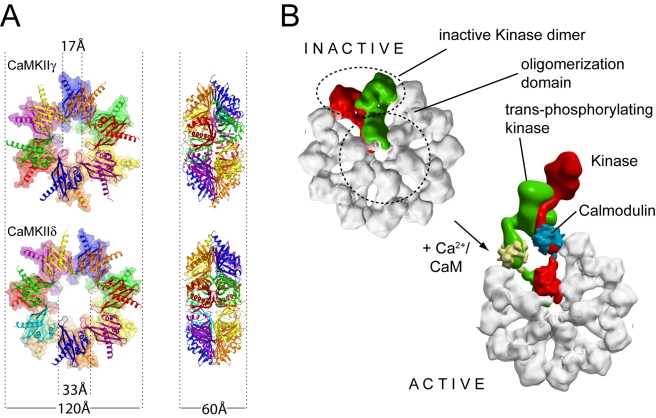
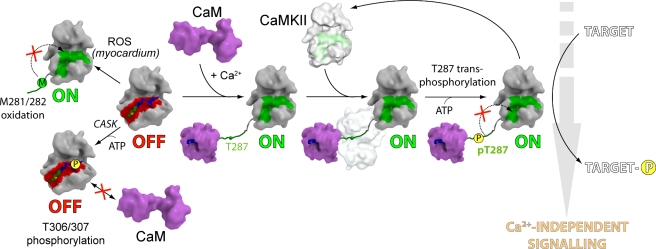
Long-term potentiation (LTP), a long-lasting enhancement in communication between neurons, is considered to be the major cellular mechanism underlying learning and memory. LTP triggers high-frequency calcium pulses that result in the activation of Calcium/Calmodulin (CaM)-dependent kinase II (CaMKII). CaMKII acts as a molecular switch because it remains active for a long time after the return to basal calcium levels, which is a unique property required for CaMKII function. Here we describe the crystal structure of the human CaMKIIdelta/Ca2+/CaM complex, structures of all four human CaMKII catalytic domains in their autoinhibited states, as well as structures of human CaMKII oligomerization domains in their tetradecameric and physiological dodecameric states. All four autoinhibited human CaMKIIs were monomeric in the determined crystal structures but associated weakly in solution. In the CaMKIIdelta/Ca2+/CaM complex, the inhibitory region adopted an extended conformation and interacted with an adjacent catalytic domain positioning T287 into the active site of the interacting protomer. Comparisons with autoinhibited CaMKII structures showed that binding of calmodulin leads to the rearrangement of residues in the active site to a conformation suitable for ATP binding and to the closure of the binding groove for the autoinhibitory helix by helix alphaD. The structural data, together with biophysical interaction studies, reveals the mechanism of CaMKII activation by calmodulin and explains many of the unique regulatory properties of these two essential signaling molecules.
Enhanced version: This article can also be viewed as an enhanced version in which the text of the article is integrated with interactive 3-D representations and animated transitions. Please note that a web plugin is required to access this enhanced functionality. Instructions for the installation and use of the Web plugin are available in Text S1.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Structure of signaling enzyme reveals how calcium turns it on.PLoS Biol. 2010 Jul 27;8(7):e1000427. doi: 10.1371/journal.pbio.1000427. PLoS Biol. 2010. PMID: 20668537 Free PMC article. No abstract available.
Similar articles
-
The KN-93 Molecule Inhibits Calcium/Calmodulin-Dependent Protein Kinase II (CaMKII) Activity by Binding to Ca2+/CaM.J Mol Biol. 2019 Mar 29;431(7):1440-1459. doi: 10.1016/j.jmb.2019.02.001. Epub 2019 Feb 10. J Mol Biol. 2019. PMID: 30753871
-
Assemblies of calcium/calmodulin-dependent kinase II with actin and their dynamic regulation by calmodulin in dendritic spines.Proc Natl Acad Sci U S A. 2019 Sep 17;116(38):18937-18942. doi: 10.1073/pnas.1911452116. Epub 2019 Aug 27. Proc Natl Acad Sci U S A. 2019. PMID: 31455737 Free PMC article.
-
Interactome Analysis Reveals Regulator of G Protein Signaling 14 (RGS14) is a Novel Calcium/Calmodulin (Ca2+/CaM) and CaM Kinase II (CaMKII) Binding Partner.J Proteome Res. 2018 Apr 6;17(4):1700-1711. doi: 10.1021/acs.jproteome.8b00027. Epub 2018 Mar 20. J Proteome Res. 2018. PMID: 29518331 Free PMC article.
-
Structural studies on the regulation of Ca2+/calmodulin dependent protein kinase II.Curr Opin Struct Biol. 2013 Apr;23(2):292-301. doi: 10.1016/j.sbi.2013.04.002. Epub 2013 Apr 27. Curr Opin Struct Biol. 2013. PMID: 23632248 Free PMC article. Review.
-
Neuronal CA2+/calmodulin-dependent protein kinase II: the role of structure and autoregulation in cellular function.Annu Rev Biochem. 2002;71:473-510. doi: 10.1146/annurev.biochem.71.110601.135410. Epub 2001 Nov 9. Annu Rev Biochem. 2002. PMID: 12045104 Review.
Cited by
-
Binding and Functional Folding (BFF): A Physiological Framework for Studying Biomolecular Interactions and Allostery.J Mol Biol. 2022 Dec 15;434(23):167872. doi: 10.1016/j.jmb.2022.167872. Epub 2022 Oct 28. J Mol Biol. 2022. PMID: 36354074 Free PMC article. Review.
-
Crystal structures of human CaMKIα reveal insights into the regulation mechanism of CaMKI.PLoS One. 2012;7(9):e44828. doi: 10.1371/journal.pone.0044828. Epub 2012 Sep 20. PLoS One. 2012. PMID: 23028635 Free PMC article.
-
PROTACs: great opportunities for academia and industry (an update from 2020 to 2021).Signal Transduct Target Ther. 2022 Jun 9;7(1):181. doi: 10.1038/s41392-022-00999-9. Signal Transduct Target Ther. 2022. PMID: 35680848 Free PMC article. Review.
-
Phosphoproteomics study based on in vivo inhibition reveals sites of calmodulin-dependent protein kinase II regulation in the heart.J Am Heart Assoc. 2013 Aug 7;2(4):e000318. doi: 10.1161/JAHA.113.000318. J Am Heart Assoc. 2013. PMID: 23926118 Free PMC article.
-
Autoinhibition and Polo-dependent multisite phosphorylation restrict activity of the histone H3 kinase Haspin to mitosis.Mol Cell. 2013 Dec 12;52(5):734-45. doi: 10.1016/j.molcel.2013.10.002. Epub 2013 Oct 31. Mol Cell. 2013. PMID: 24184212 Free PMC article.
References
-
- Manning G, Whyte D. B, Martinez R, Hunter T, Sudarsanam S. The protein kinase complement of the human genome. Science. 2002;298:1912–1934. - PubMed
-
- Hudmon A, Schulman H. Neuronal CA2+/calmodulin-dependent protein kinase II: the role of structure and autoregulation in cellular function. Annu Rev Biochem. 2002;71:473–510. - PubMed
-
- Tobimatsu T, Fujisawa H. Tissue-specific expression of four types of rat calmodulin-dependent protein kinase II mRNAs. J Biol Chem. 1989;264:17907–17912. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
