

Innate immune suppression enables frequent transfection with RNA encoding reprogramming proteins
- PMID: 20668695
- PMCID: PMC2909252
- DOI: 10.1371/journal.pone.0011756
Innate immune suppression enables frequent transfection with RNA encoding reprogramming proteins
Abstract
Background: Generating autologous pluripotent stem cells for therapeutic applications will require the development of efficient DNA-free reprogramming techniques. Transfecting cells with in vitro-transcribed, protein-encoding RNA is a straightforward method of directly expressing high levels of reprogramming proteins without genetic modification. However, long-RNA transfection triggers a potent innate immune response characterized by growth inhibition and the production of inflammatory cytokines. As a result, repeated transfection with protein-encoding RNA causes cell death.
Methodology/principal findings: RNA viruses have evolved methods of disrupting innate immune signaling by destroying or inhibiting specific proteins to enable persistent infection. Starting from a list of known viral targets, we performed a combinatorial screen to identify siRNA cocktails that could desensitize cells to exogenous RNA. We show that combined knockdown of interferon-beta (Ifnb1), Eif2ak2, and Stat2 rescues cells from the innate immune response triggered by frequent long-RNA transfection. Using this technique, we were able to transfect primary human fibroblasts every 24 hours with RNA encoding the reprogramming proteins Oct4, Sox2, Klf4, and Utf1. We provide evidence that the encoded protein is active, and we show that expression can be maintained for many days, through multiple rounds of cell division.
Conclusions/significance: Our results demonstrate that suppressing innate immunity enables frequent transfection with protein-encoding RNA. This technique represents a versatile tool for investigating expression dynamics and protein interactions by enabling precise control over levels and timing of protein expression. Our finding also opens the door for the development of reprogramming and directed-differentiation methods based on long-RNA transfection.
Conflict of interest statement
Figures
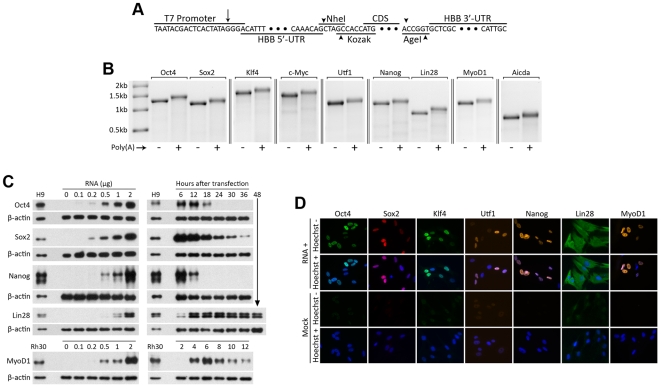
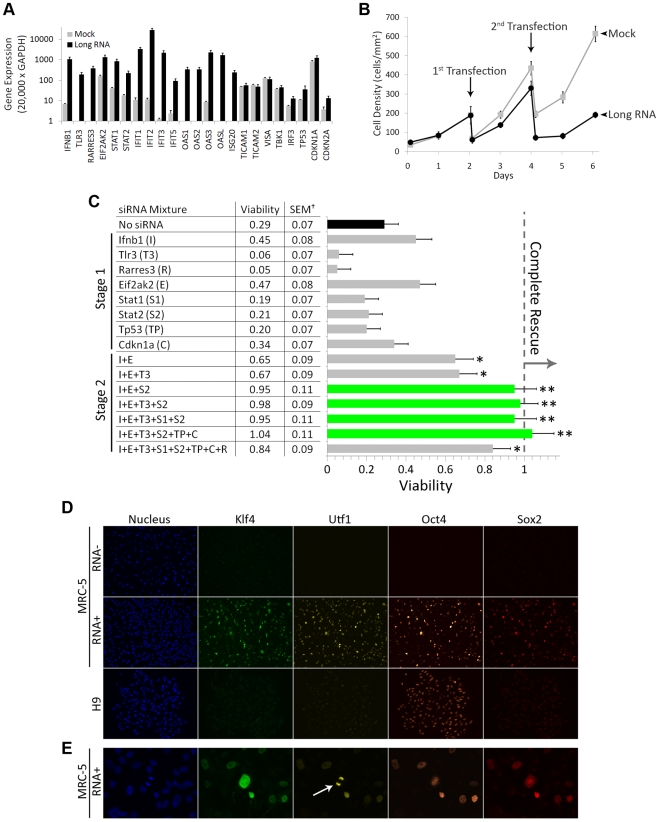
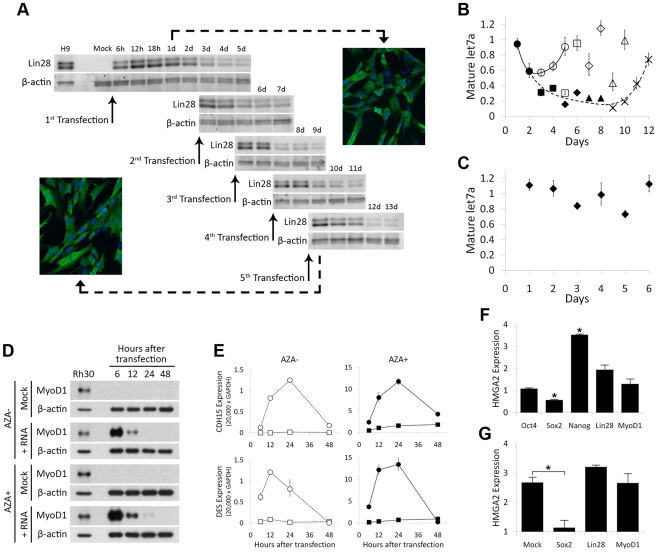
References
-
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. - PubMed
-
- Okita K, Ichisaka T, Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007;448:313–317. - PubMed
-
- Wernig M, Meissner A, Foreman R, Brambrink T, Ku M, et al. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature. 2007;448:318–324. - PubMed
-
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. - PubMed
-
- Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, et al. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318:1917–1920. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
