

Strong reproductive barriers in a narrow hybrid zone of West-Mediterranean green toads (Bufo viridis subgroup) with Plio-Pleistocene divergence
- PMID: 20670415
- PMCID: PMC2923517
- DOI: 10.1186/1471-2148-10-232
Strong reproductive barriers in a narrow hybrid zone of West-Mediterranean green toads (Bufo viridis subgroup) with Plio-Pleistocene divergence
Abstract
Background: One key question in evolutionary biology deals with the mode and rate at which reproductive isolation accumulates during allopatric speciation. Little is known about secondary contacts of recently diverged anuran species. Here we conduct a multi-locus field study to investigate a contact zone between two lineages of green toads with an estimated divergence time of 2.7 My, and report results from preliminary experimental crosses.
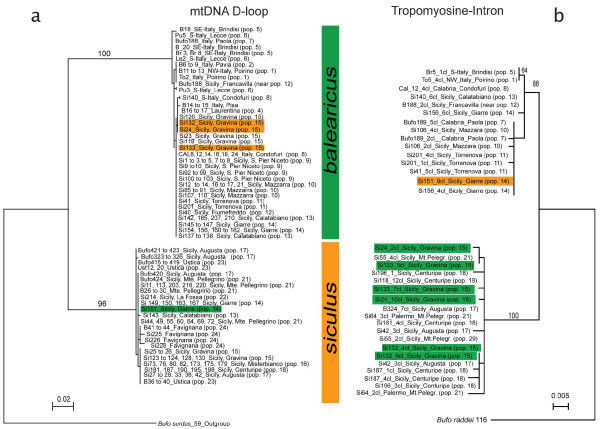
Results: The Sicilian endemic Bufo siculus and the Italian mainland-origin B. balearicus form a narrow hybrid zone east of Mt. Etna. Despite bidirectional mtDNA introgression over a ca. 40 km North-South cline, no F1 hybrids could be found, and nuclear genomes display almost no admixture. Populations from each side of the contact zone showed depressed genetic diversity and very strong differentiation (FST = 0.52). Preliminary experimental crosses point to a slightly reduced fitness in F1 hybrids, a strong hybrid breakdown in backcrossed offspring (F1 x parental, with very few reaching metamorphosis) and a complete and early mortality in F2 (F1 x F1).
Conclusion: Genetic patterns at the contact zone are molded by drift and selection. Local effective sizes are reduced by the geography and history of the contact zone, B. balearicus populations being at the front wave of a recent expansion (late Pleistocene). Selection against hybrids likely results from intrinsic genomic causes (disruption of coadapted sets of genes in backcrosses and F2-hybrids), possibly reinforced by local adaptation (the ranges of the two taxa roughly coincide with the borders of semiarid and arid climates). The absence of F1 in the field might be due to premating isolation mechanisms. Our results, show that these lineages have evolved almost complete reproductive isolation after some 2.7 My of divergence, contrasting sharply with evidence from laboratory experiments that some anuran species may still produce viable F1 offspring after > 20 My of divergence.
Figures






References
-
- Coyne JA, Orr HA. Speciation. Sunderland, Sinauer, MA; 2004.
-
- Johns GC, Avise JC. A comparative summary of genetic distances in the vertebrates from the mitochondrial cytochrome b gene. Mol Biol Evol. 1998;15(11):1481–1490. - PubMed
-
- Blair WF. Evolution in the genus Bufo. Austin and London; 1972.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
