

The structure and function of serum opacity factor: a unique streptococcal virulence determinant that targets high-density lipoproteins
- PMID: 20671930
- PMCID: PMC2910554
- DOI: 10.1155/2010/956071
The structure and function of serum opacity factor: a unique streptococcal virulence determinant that targets high-density lipoproteins
Abstract
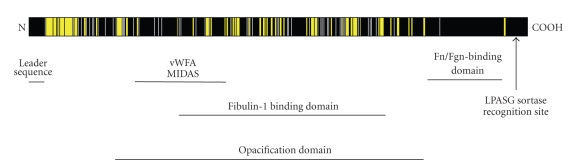
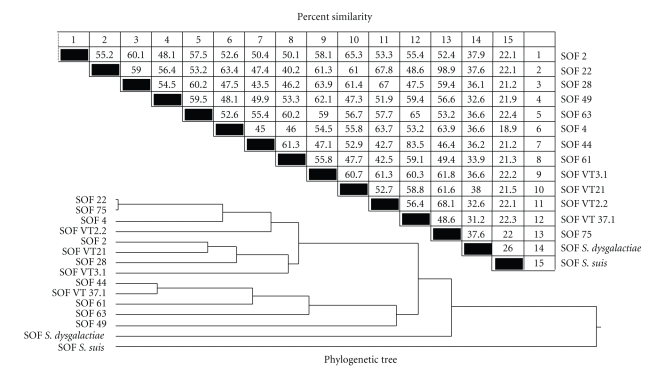
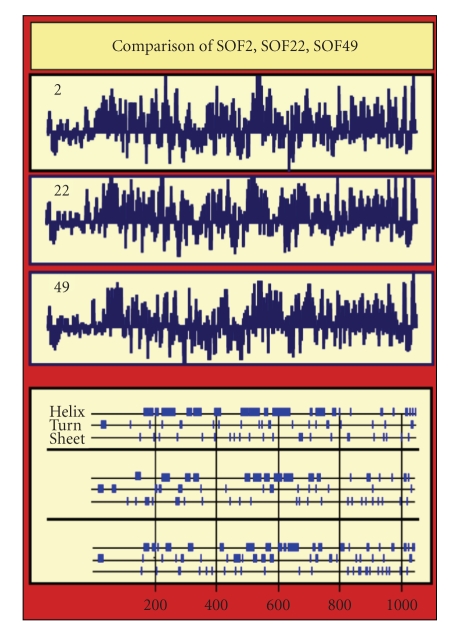
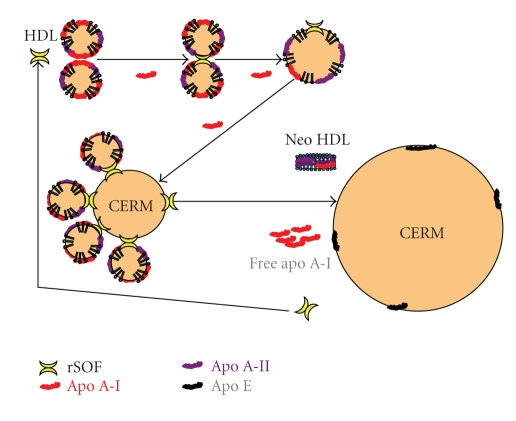
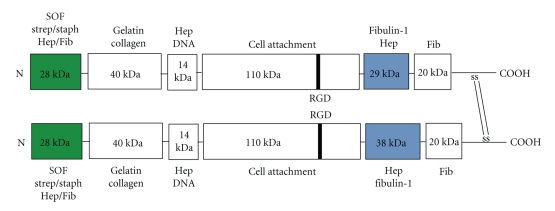
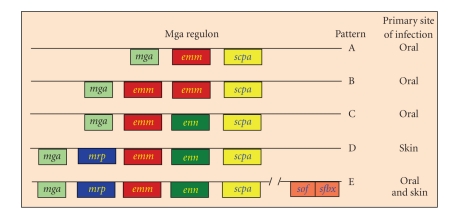
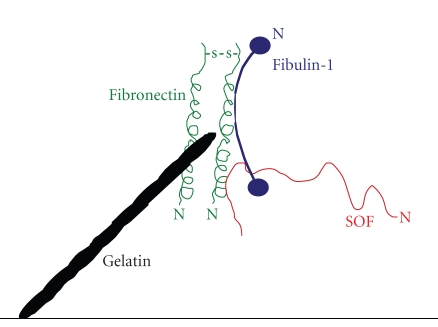
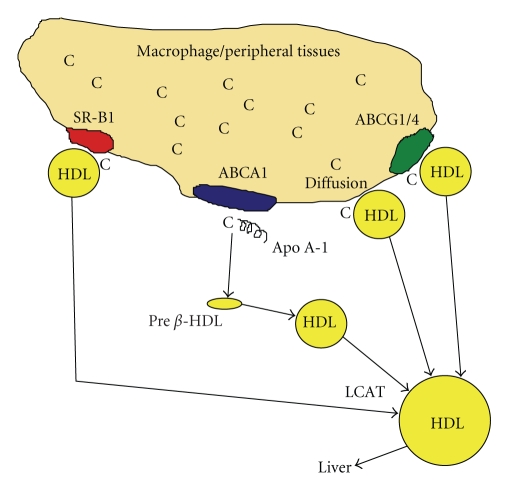
Serum opacity factor (SOF) is a virulence determinant expressed by a variety of streptococcal and staphylococcal species including both human and animal pathogens. SOF derives its name from its ability to opacify serum where it targets and disrupts the structure of high-density lipoproteins resulting in formation of large lipid vesicles that cause the serum to become cloudy. SOF is a multifunctional protein and in addition to its opacification activity, it binds to a number of host proteins that mediate adhesion of streptococci to host cells, and it plays a role in resistance to phagocytosis in human blood. This article will provide an overview of the structure and function of SOF, its role in the pathogenesis of streptococcal infections, its vaccine potential, its prevalence and distribution in bacteria, and the molecular mechanism whereby SOF opacifies serum and how an understanding of this mechanism may lead to therapies for reducing high-cholesterol concentrations in blood, a major risk factor for cardiovascular disease.
Figures


References
-
- Ward HK, Rudd GV. Studies on haemolytic streptococci from human sources. Australian Journal of Experimental Biology & Medical Science. 1938;16:181–192.
-
- Rowen R, Martin J. Enhancement of cholesterol esterification in serum by an extract of group-A streptococcus. Biochimica et Biophysica Acta. 1963;70:396–405. - PubMed
-
- Saravani GA, Martin DR. Opacity factor from group A streptococci is an apoproteinase. FEMS Microbiology Letters. 1990;68(1-2):35–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
