

Review
doi: 10.1002/cm.20473.
Conformational dynamics of actin: effectors and implications for biological function
Affiliations
- PMID: 20672362
- PMCID: PMC3038201
- DOI: 10.1002/cm.20473
Item in Clipboard
Review
Conformational dynamics of actin: effectors and implications for biological function
Cytoskeleton (Hoboken).
2010 Oct.
Free PMC article
Abstract
Actin is a protein abundant in many cell types. Decades of investigations have provided evidence that it has many functions in living cells. The diverse morphology and dynamics of actin structures adapted to versatile cellular functions is established by a large repertoire of actin-binding proteins. The proper interactions with these proteins assume effective molecular adaptations from actin, in which its conformational transitions play essential role. This review attempts to summarise our current knowledge regarding the coupling between the conformational states of actin and its biological function.
Figures
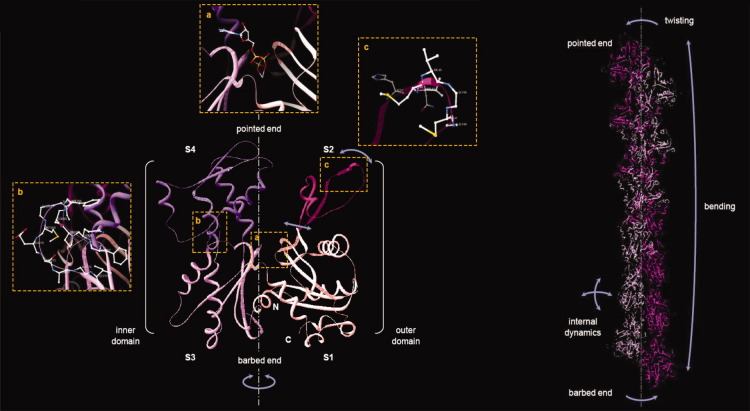
(A) Crystal structure of the G-actin molecule. Actin subdomains are indicated by different colors and numbers according to Holmes et al (S1) amino acids 1–32, 70–144, 338–375; (S2) amino acids 33–69; (S3) amino acids 145–180, 270–337; (S4) amino acids 181–269 [Holmes et al., 1990]. The amino-, and carboxyl termini of the molecule are indicated by N and C, respectively. Orange-dotted boxes depict enlarged views of the nucleotide binding cleft with bound ATP (a), the structure of the hydrophobic loop (b, amino acids 262–274, rotated by 90° to the right with respect to the main axis of the monomer, shown by gray dashed-dotted line) and the DNase binding loop (c, amino acids 40–48). The schematic representation of the conformational dynamics on the basis of the normal mode analysis of G-actin is shown by blue arrows [Tirion and ben-Avraham, 1993]. The image was made on the basis of the crystal structure of rabbit skeletal muscle actin in complex with DNase I (PDB code 1ATN, [Kabsch et al., 1990]). (B) Helical organization of actin protomers in the filament. A structural model of a 13-mer F-actin was derived from Oda's F-actin protomer model (PDB code 2ZWH, [Oda et al., 2009]). The two linear strands of actin protomers composing the two-start right-handed long pitch helix are colored by dim and dark violet, respectively. The single-start left-handed short pith helix is assembled by alternating dim and dark actin protomers. The schematic representation of the conformational dynamics of F-actin is shown by blue arrows. The ribbon diagrams were obtained with Deep View/Swiss PDB Viewer.
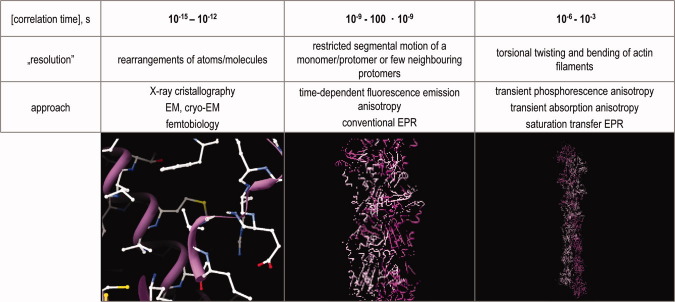
The table shows the corresponding correlation times and the suitable approaches for their investigation.
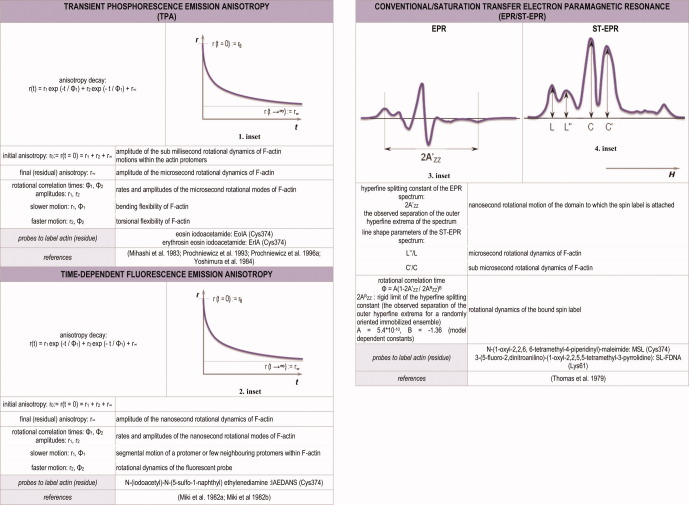
The formulation and parameters of transient phosphorescence emission anisotropy (TPA), time-dependent fluorescence emission anisotropy and conventional/saturation transfer (ST) EPR. Typical phosphorescence (1. inset)/fluorescence (2. inset) anisotropy decay (r(t)) of actin, and typical EPR (3. inset)/ST-EPR (4. inset) spectra of actin are shown. H is releated to the direction and the strength of the applied magnetic field. In phosphorescence/fluorescence emission anisotropy the kinetics of anisotropy decay, while in EPR/ST-EPR the shape of the spectrum characteristic for the conformational dynamics of the molecule. [Color figure can be viewed in the online issue which is available at http://wileyonlinelibrary.com .]
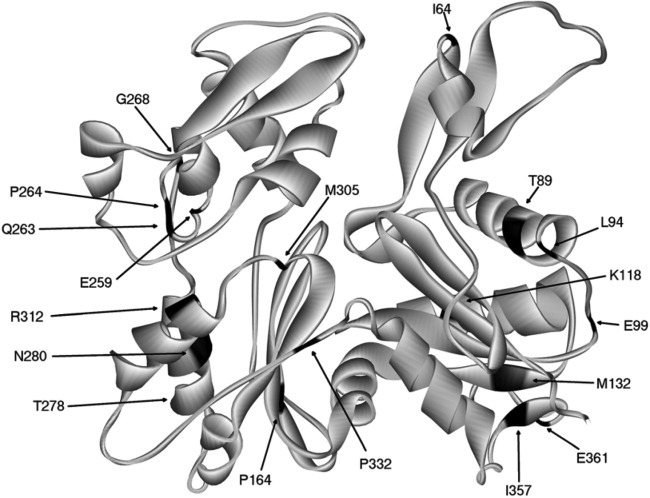
The most common mutation sites are labeled with one letter and ID number of the amino acids [Olson et al., ; van Wijk et al., ; Zhu et al., ; Ilkovski et al., ; Marston et al., 2004]. The ribbon structure of rabbit skeletal muscle actin (PDB code: 1ATN [Kabsch et al., 1990] is colored black at the mutated points.
Similar articles
-
The actin cytoskeleton of Dictyostelium: a story told by mutants.J Cell Sci. 2000 Mar;113 ( Pt 5):759-66. doi: 10.1242/jcs.113.5.759. J Cell Sci. 2000. PMID: 10671366 Review.
-
Regulation of the cytoskeleton assembly: a role for a ternary complex of actin with two actin-binding proteins.Results Probl Cell Differ. 2001;32:165-79. doi: 10.1007/978-3-540-46560-7_12. Results Probl Cell Differ. 2001. PMID: 11131830 Review. No abstract available.
-
Regulation of actin cytoskeleton dynamics in cells.Mol Cells. 2010 Apr;29(4):311-25. doi: 10.1007/s10059-010-0053-8. Mol Cells. 2010. PMID: 20446344 Free PMC article. Review.
-
Signaling to the actin cytoskeleton.Annu Rev Cell Dev Biol. 1998;14:305-38. doi: 10.1146/annurev.cellbio.14.1.305. Annu Rev Cell Dev Biol. 1998. PMID: 9891786 Review.
-
Cytoskeleton: actin and endocytosis--no longer the weakest link.Curr Biol. 2001 Sep 4;11(17):R691-4. doi: 10.1016/s0960-9822(01)00410-9. Curr Biol. 2001. PMID: 11553341 Review.
Cited by
-
MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation.Mol Cell. 2013 Aug 8;51(3):397-404. doi: 10.1016/j.molcel.2013.06.019. Epub 2013 Aug 1. Mol Cell. 2013. PMID: 23911929 Free PMC article.
-
Mitochondrial dysfunction and mitochondrial dynamics-The cancer connection.Biochim Biophys Acta Bioenerg. 2017 Aug;1858(8):602-614. doi: 10.1016/j.bbabio.2017.01.004. Epub 2017 Jan 16. Biochim Biophys Acta Bioenerg. 2017. PMID: 28104365 Free PMC article. Review.
-
Myosin and tropomyosin stabilize the conformation of formin-nucleated actin filaments.J Biol Chem. 2012 Sep 14;287(38):31894-904. doi: 10.1074/jbc.M112.341230. Epub 2012 Jun 29. J Biol Chem. 2012. PMID: 22753415 Free PMC article.
-
Actin polymerization negatively regulates p53 function by impairing its nuclear import in response to DNA damage.PLoS One. 2013;8(4):e60179. doi: 10.1371/journal.pone.0060179. Epub 2013 Apr 2. PLoS One. 2013. PMID: 23565200 Free PMC article.
-
Ethanol Effect on BK Channels is Modulated by Magnesium.Alcohol Clin Exp Res. 2015 Sep;39(9):1671-9. doi: 10.1111/acer.12821. Alcohol Clin Exp Res. 2015. PMID: 26331878 Free PMC article.
References
-
- Allen PG, Laham LE, Way M, Janmey PA. Binding of phosphate, aluminum fluoride, or beryllium fluoride to F-actin inhibits severing by gelsolin. J Biol Chem. 1996;271:4665–4670. - PubMed
-
- Andrianantoandro E, Pollard TD. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol Cell. 2006;24:13–23. - PubMed
-
- Arii Y, Hatori K. Relationship between the flexibility and the motility of actin filaments: effects of pH. Biochem Biophys Res Commun. 2008;371:772–776. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
