

Regenerative therapy and cancer: in vitro and in vivo studies of the interaction between adipose-derived stem cells and breast cancer cells from clinical isolates
- PMID: 20673000
- PMCID: PMC3011910
- DOI: 10.1089/ten.TEA.2010.0248
Regenerative therapy and cancer: in vitro and in vivo studies of the interaction between adipose-derived stem cells and breast cancer cells from clinical isolates
Abstract
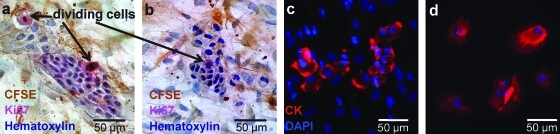
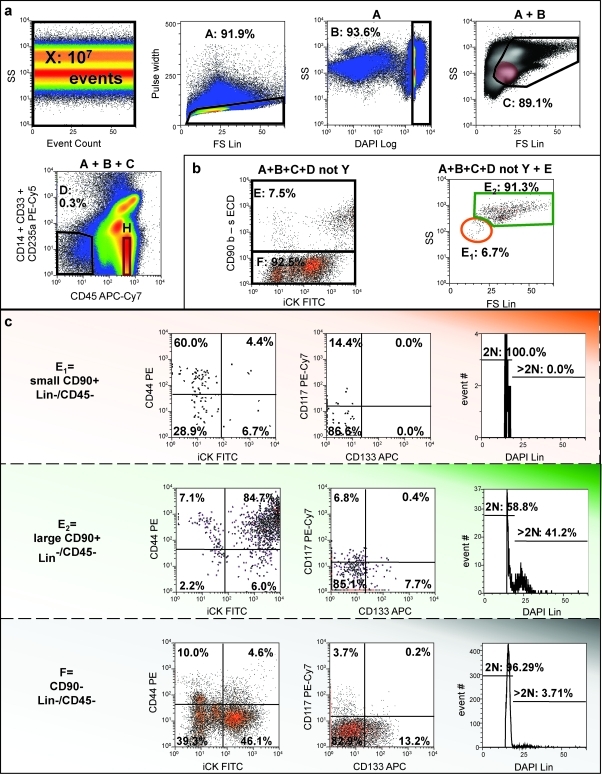
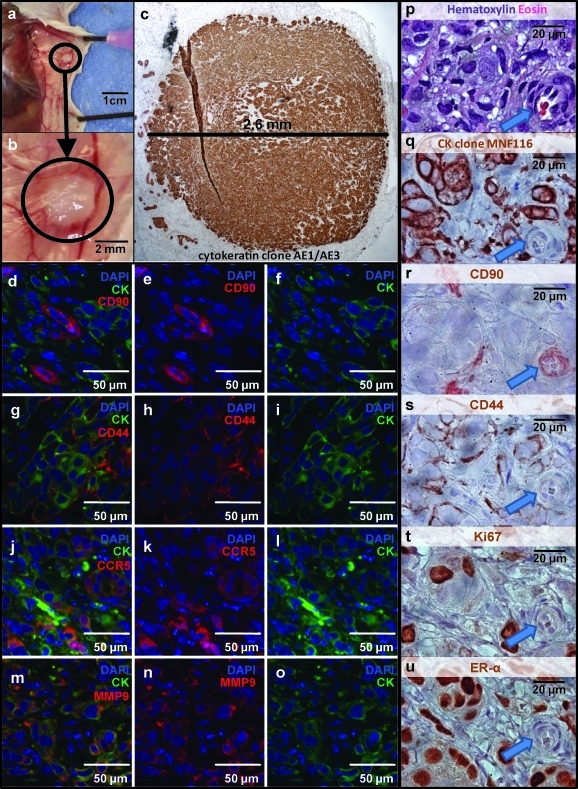
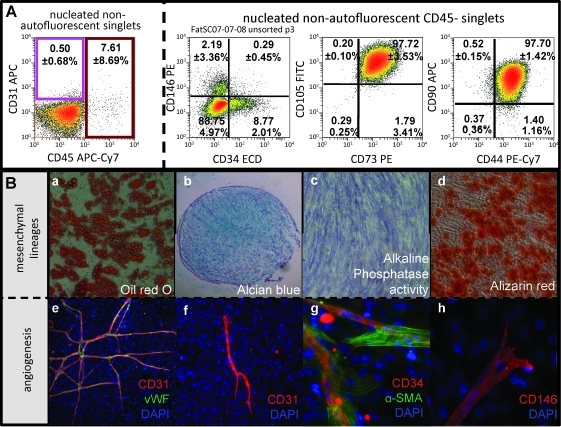
Adipose-derived stem cells (ASCs) have been proposed to stabilize autologous fat grafts for regenerative therapy, but their safety is unknown in the setting of reconstructive surgery after mastectomy. Both bone marrow mesenchymal stem cells (MSCs) and ASC have been shown to enhance tumorigenesis of established breast cancer cell lines, but primary patient material has not been tested. Here, we ask whether ASC promote the in vitro growth and in vivo tumorigenesis of metastatic breast cancer clinical isolates. Metastatic pleural effusion (MPE) cells were used for coculture experiments. ASC enhanced the proliferation of MPE cells in vitro (5.1-fold). For xenograft experiments (100 sorted cells/injection site), nonhematopoietic MPE cells were sorted into resting and active populations: CD90+ resting (low scatter, 2.1%≥2N DNA), CD90+ active (high scatter, 10.6%≥2N DNA), and CD90-. Resting CD90+ MPE cells were tumorigenic in 4/40 sites but growth was not augmented by ASC. Active CD90+ MPE cells were tumorigenic (17/40 sites) only when coinjected with ASC (p=0.0005, χ2 test). The multilineage potentiality and MSC-like immunophenotype of ASC were confirmed by flow cytometry, differentiation cultures, and immunostaining. The secretome profile of ASC resembled that reported for MSC, but included adipose-associated adipsin and the hormone leptin, shown to promote breast cancer growth. Our data indicate that ASC enhance the growth of active, but not resting tumor cells. Thus, reconstructive therapy utilizing ASC-augmented whole fat should be postponed until there is no evidence of active disease.
Figures
Similar articles
-
Adipose-derived stem cell-based treatment for acute liver failure.Stem Cell Res Ther. 2015 Mar 21;6(1):40. doi: 10.1186/s13287-015-0040-2. Stem Cell Res Ther. 2015. PMID: 25890008 Free PMC article.
-
Amniotic Mesenchymal Stromal Cells Exhibit Preferential Osteogenic and Chondrogenic Differentiation and Enhanced Matrix Production Compared With Adipose Mesenchymal Stromal Cells.Am J Sports Med. 2017 Sep;45(11):2637-2646. doi: 10.1177/0363546517706138. Epub 2017 May 25. Am J Sports Med. 2017. PMID: 28541092 Free PMC article.
-
Adipose tissue-derived mesenchymal stem cell yield and growth characteristics are affected by the tissue-harvesting procedure.Cytotherapy. 2006;8(2):166-77. doi: 10.1080/14653240600621125. Cytotherapy. 2006. PMID: 16698690
-
The Crosstalk Between Adipose-Derived Stem or Stromal Cells (ASC) and Cancer Cells and ASC-Mediated Effects on Cancer Formation and Progression-ASCs: Safety Hazard or Harmless Source of Tropism?Stem Cells Transl Med. 2022 Apr 29;11(4):394-406. doi: 10.1093/stcltm/szac002. Stem Cells Transl Med. 2022. PMID: 35274703 Free PMC article. Review.
-
Adipose-derived stem cells in cartilage regeneration: current perspectives.Regen Med. 2016 Oct;11(7):693-703. doi: 10.2217/rme-2016-0077. Epub 2016 Sep 6. Regen Med. 2016. PMID: 27599358 Review.
Cited by
-
Adipose-Derived Stromal Vascular Fraction Differentially Expands Breast Progenitors in Tissue Adjacent to Tumors Compared to Healthy Breast Tissue.Plast Reconstr Surg. 2015 Oct;136(4):414e-425e. doi: 10.1097/PRS.0000000000001635. Plast Reconstr Surg. 2015. PMID: 26090768 Free PMC article.
-
Roles of Matrix Metalloproteinases and Their Natural Inhibitors in Metabolism: Insights into Health and Disease.Int J Mol Sci. 2023 Jun 26;24(13):10649. doi: 10.3390/ijms241310649. Int J Mol Sci. 2023. PMID: 37445827 Free PMC article. Review.
-
Stem cell treatment for patients with autoimmune disease by systemic infusion of culture-expanded autologous adipose tissue derived mesenchymal stem cells.J Transl Med. 2011 Oct 21;9:181. doi: 10.1186/1479-5876-9-181. J Transl Med. 2011. PMID: 22017805 Free PMC article.
-
Immunohistological localization of endogenous unlabeled stem cells in wounded skin.J Histochem Cytochem. 2014 Apr;62(4):276-85. doi: 10.1369/0022155414520710. Epub 2014 Jan 7. J Histochem Cytochem. 2014. PMID: 24399040 Free PMC article.
-
Alterations of gene expression and protein synthesis in co-cultured adipose tissue-derived stem cells and squamous cell-carcinoma cells: consequences for clinical applications.Stem Cell Res Ther. 2014 May 12;5(3):65. doi: 10.1186/scrt454. Stem Cell Res Ther. 2014. PMID: 24887580 Free PMC article.
References
-
- Neuber G.A. Fettransplantation. Chir Kongr Verhandl Deutsche Gesellschaft für Chirurgie. 1893;22:66.
-
- Czerny V. Plastischer Ersatz der Brustdruse durch ein Lipom. Chir Kong Verhandl. 1895;2:216.
-
- ASPRS. Report on autologous fat transplantation. ASPRS Ad-Hoc Committee on New Procedures, September 30, 1987. Plast Surg Nurs. 1987;7:140. - PubMed
-
- Delay E. Garson S. Tousson G. Sinna R. Fat injection to the breast: technique, results, and indications based on 880 procedures over 10 years. Aesthet Surg J. 2009;29:360. - PubMed
-
- Peer L.A. Loss of Weight and Volume in Human Fat Grafts: With Postulation of A “Cell Survival Theory.”. Plast Reconstr Surg. 1950;5:217.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
