

Some considerations for analyzing biodiversity using integrative metagenomics and gene networks
- PMID: 20673351
- PMCID: PMC2921367
- DOI: 10.1186/1745-6150-5-47
Some considerations for analyzing biodiversity using integrative metagenomics and gene networks
Abstract
Background: Improving knowledge of biodiversity will benefit conservation biology, enhance bioremediation studies, and could lead to new medical treatments. However there is no standard approach to estimate and to compare the diversity of different environments, or to study its past, and possibly, future evolution.
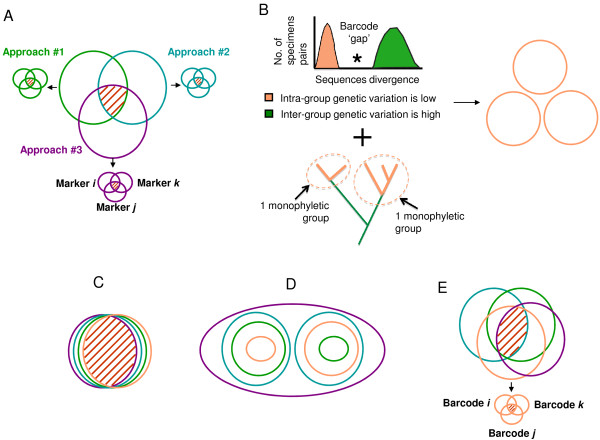
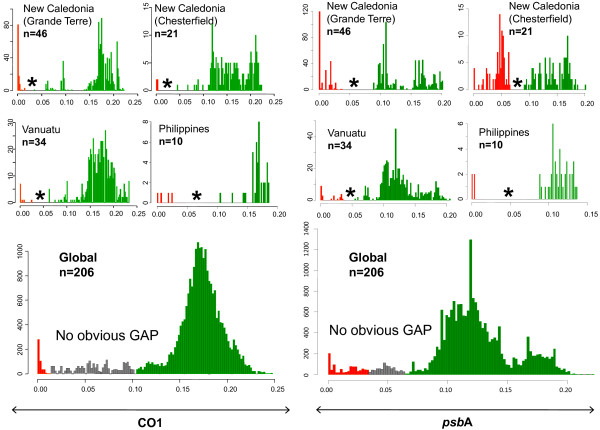
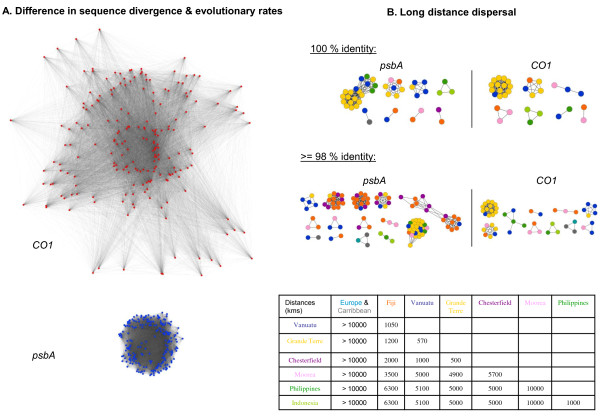
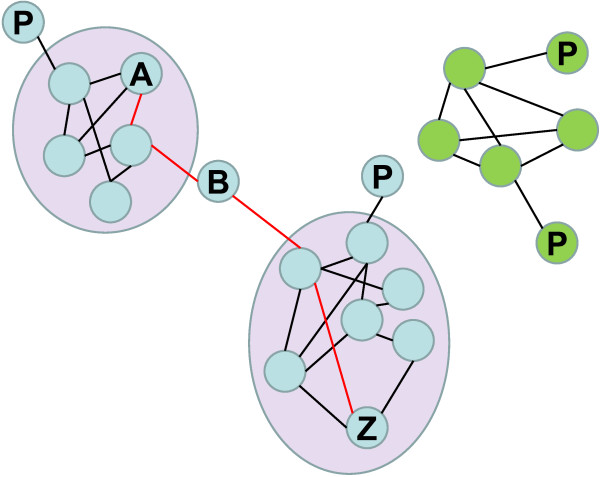
Presentation of the hypothesis: We argue that there are two conditions for significant progress in the identification and quantification of biodiversity. First, integrative metagenomic studies - aiming at the simultaneous examination (or even better at the integration) of observations about the elements, functions and evolutionary processes captured by the massive sequencing of multiple markers - should be preferred over DNA barcoding projects and over metagenomic projects based on a single marker. Second, such metagenomic data should be studied with novel inclusive network-based approaches, designed to draw inferences both on the many units and on the many processes present in the environments.
Testing the hypothesis: We reached these conclusions through a comparison of the theoretical foundations of two molecular approaches seeking to assess biodiversity: metagenomics (mostly used on prokaryotes and protists) and DNA barcoding (mostly used on multicellular eukaryotes), and by pragmatic considerations of the issues caused by the 'species problem' in biodiversity studies.
Implications of the hypothesis: Evolutionary gene networks reduce the risk of producing biodiversity estimates with limited explanatory power, biased either by unequal rates of LGT, or difficult to interpret due to (practical) problems caused by type I and type II grey zones. Moreover, these networks would easily accommodate additional (meta)transcriptomic and (meta)proteomic data.
Figures
References
-
- Morgan GJ. Evaluating Maclaurin and Sterelny's Conception of Biodiversity in Cases of Frequent, Promiscuous Lateral Gene Transfer. Biology and Philosophy. 2010. in press .
-
- Brennerova MV, Josefiova J, Brenner V, Pieper DH, Junca H. Metagenomics reveals diversity and abundance of meta-cleavage pathways in microbial communities from soil highly contaminated with jet fuel under air-sparging bioremediation. Environ Microbiol. 2009;11:2216–2227. doi: 10.1111/j.1462-2920.2009.01943.x. - DOI - PMC - PubMed
-
- Wolcott RD, Gontcharova V, Sun Y, Dowd SE. Evaluation of the bacterial diversity among and within individual venous leg ulcers using bacterial tag-encoded FLX and titanium amplicon pyrosequencing and metagenomic approaches. BMC Microbiol. 2009;9:226. doi: 10.1186/1471-2180-9-226. - DOI - PMC - PubMed
-
- Doolittle WF, Zhaxybayeva O. Metagenomics and the Units of Biological Organization. Bioscience. 2010;60:102–112. doi: 10.1525/bio.2010.60.2.5. - DOI
-
- Callicott JB, Crowder LB, Mumford K. Current normative concepts in conservation. Conservation Biology. 1999;13:22–35. doi: 10.1046/j.1523-1739.1999.97333.x. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
